
神经胶质细胞
| 神经胶质细胞 | |
|---|---|
 | |
| 基本信息 | |
| 发育自 | Neuroectoderm for macroglia, and hematopoietic stem cells for microglia |
| 系统 | 神经系统 |
| 标识字符 | |
| MeSH | D009457 |
| TA98 | A14.0.00.005 |
| TH | H2.00.06.2.00001 |
| FMA | FMA:54541、54536 |
| 《显微解剖学术语》 [在维基数据上编辑] | |

神经胶质细胞(neurogliocyte[1][2])是中枢神经系统和周围神经系统中的“非神经元细胞”,不会产生电脉冲。[3] 它们种类繁多,功能是维持体内稳态、为神经元提供支持和保护、并可在周围神经形成髓鞘。[4] 神经胶质细胞的汉英异名均庞杂,因此又称神经胶细胞(neuroglial cell[5])、胶质细胞(gliocyte)、胶细胞[6](glial cell)、神经胶质(neuroglia)、神经胶[7]、简称胶质[8](glia),
在中枢神经系统中,神经胶质细胞包括寡突胶质细胞、星形胶质细胞、室管膜细胞和小胶质细胞;在周围神经系统中,神经胶质细胞包括施旺细胞和卫星细胞。 它们具有四个主要功能:(1)围绕神经元并将其固定在适当位置; (2)为神经元提供营养和氧; (3)使一个神经元与另一个神经元绝缘; (4)消灭病原体并清除死亡的神经元。 它们还在神经传递、突触连接以及呼吸等生理过程中发挥作用。[9][10][11][12]
虽然人们认为神经胶质细胞比神经元的比例为10:1,但最近的一项研究提供了低于1:1的比例的证据。[13] 此外,神经胶质细胞比神经元具有更多的细胞多样性和功能,并且神经胶质细胞可以多种方式响应和操纵神经传导。 此外,它们可以影响记忆的保存和巩固。[3]
神经胶质细胞是病理学家鲁道夫·菲尔绍在寻找脑中的结缔组织时于1856年发现的。[14] 该术语源自希腊语 γλία 和 γλοία (胶),暗示了它们最初被认为是神经系统中的胶水。
分类
[编辑]大胶质细胞
[编辑]| 存在位置 | 名称 | 描述 |
|---|---|---|
| 中枢 | 星形胶质细胞 | 星形胶质细胞,又称星状细胞,为胶质细胞中体积最大的一种,细胞核较大、浅染,呈圆形或卵圆形,细胞体发出许多胞突,胞突伸展填充在神经元胞体及突起之间,可支持和固定作用。部分胞突末端膨大为脚板。脚板贴附于毛细血管壁上,构成脑血管障壁;附着在脑和脊髓表面,形成胶质界膜(glial limitans)。根据星形胶质细胞胞突的多寡分为两类:纤维性星形胶质细胞的胞突长、分支少,主要出现于白质里;原浆性星形胶质细胞的胞突短、分支多,主要出现于灰质里;两者功能相似。
它们透过移除过剩的离子(特别是钾离子)并回收在突触传递中放出的神经传导物,以调控神经元的外部化学环境。星状细胞可分泌花生四烯酸,而其代谢物为一具活性、可调控血管舒张及血管收缩。星形胶质细胞乃透过钙来互相传递讯息。星状细胞因彼此之间有隙型连接(亦称为电性突触),因此讯息分子,例如肌醇三磷酸(inositol triphosphate, IP3),可以自由地自一个星状细胞扩散到另一个细胞。IP3引发的传讯路径会开启细胞内胞器上的钙离子通道,让钙离子进入细胞质。这些钙离子可能再促发更多IP3的产生。这个反应的净结果为创造一股钙离子波在细胞间传递。星状细胞于胞外释放三磷酸腺苷(ATP),并活化其他星状细胞的嘌呤受体,在某些情况下也可能产生此种钙离子波。 |
| 中枢 | 寡突胶质细胞 | 寡突胶质细胞体积较星形胶质细胞小,细胞核呈卵圆形,染色质致密,细胞质内含较多微管。在银染标本中,寡突胶质细胞突起少,但在免疫细胞化学染色上可见突起较多,分支也较多。寡突胶质细胞的突起末端扩展成扁平薄膜,包覆中枢神经系统中部分神经元的轴突,形成髓鞘。髓鞘可提供绝缘效果,使神经讯号传递得更快、更有效率。 |
| 中枢 | 室管膜细胞 | 室管膜细胞呈立方或柱状,表面有很多微绒毛,部分细胞表面有纤毛;有些细胞在基底侧长有一条细长突起伸向深部,称为伸长细胞(tanycyte)。主要分布于脑室内壁,形成单层上皮,称为室管膜(ependyma);可制造并分泌脑脊液,并借由细胞上的纤毛帮助脑脊液的循环。 |
| 中枢 | 放射状胶质细胞 | 放射状胶质细胞在神经发生的早期即自神经上皮细胞衍生。此种细胞的分化能力比神经上皮细胞更为受限。在发育中的神经系统中,放射状胶质细胞一方面是神经元的前身,一方面也充当新生神经元迁移的支架。在成熟的脑中,小脑和视网膜仍保留有典型的放射状胶质细胞。在小脑中,此种细胞称为伯格曼胶质细胞(Bergmann glia),可调控突触可塑性。在视网膜中,放射状的米勒细胞(Müller cell)为主要的胶质细胞,并参与神经元间的双向沟通。 |
| 周围 | 许旺细胞 | 许旺细胞(又名神经膜细胞)在周围神经系统中形成髓鞘,与寡突胶质细胞功能相似。此种细胞亦具有吞噬能力,可清除细胞残渣,提供周围神经系统的神经元重生空间。 |
| 周围 | 卫星细胞 | 卫星细胞在神经节内包裹神经元胞体的一层扁平或立方细胞,又名被囊细胞(capsular cell)。负责调节神经元的外部化学环境。 |
小胶质细胞
[编辑]小胶质细胞是胶质细胞中最小的一种,在中枢神经系统中大约占总细胞数的15%,在脑及脊髓中各处可见。它的细胞核小、深染,呈扁平状或三角形,胞体发出细长有分支的突起,突起表面还有很多小的棘突。小胶质细胞为特化的巨噬细胞,在中枢神经系统中进行吞噬作用,清除病原体或受损的神经元。虽然小胶质细胞是由造血前驱细胞衍生而来,而非外胚层组织的一部分,因此严格说来不算胶质细胞,但因其在神经系统中支持性的角色,通常仍将其归为胶质细胞。
其他
[编辑]脑垂体后叶的垂体细胞是一种神经胶质细胞,与星形胶质细胞类似。[15] 下丘脑正中隆起的伸长细胞是一种室管膜细胞,从放射神经胶质细胞下降排列在第三脑室的底部。[16]
总数量
[编辑]通常,神经胶质细胞的体积比神经元小。 从前认为人脑中的神经胶质细胞数量接近一万亿,与神经元的数量比为10:1,但有新的证据显示人脑中大约只有850亿个神经胶质细胞,与神经元的数目大致相同。[13] [17]
发育
[编辑]大部分的胶质细胞自发育中胚胎的外胚层组织衍生而来,特别是神经管及神经脊;唯一例外者为自造血干细胞衍生而来的小胶质细胞。在成人的身体中,小胶质细胞为可自我更新的一个族群,与中枢神经系统受损时会渗入的巨噬细胞及单核细胞有明显不同。
在中枢神经系统,胶质细胞发育自神经管的脑室区(ventricular zone);此类细胞包括少突胶质细胞、室管膜细胞、星形胶质细胞。在周围神经系统,胶质胞发育自神经脊;此类细胞包括神经中的许旺细胞及神经节中的卫星细胞。
功能
[编辑]一些神经胶质细胞主要充当神经元的物理支持。 其他的则为神经元提供营养,并调节大脑的细胞外液,尤其是周围的神经元及其突触。 在早期胚胎发生期间,神经胶质细胞指导神经元的迁移并产生修饰轴突和树突生长的分子。
神经元修复与发育
[编辑]神经胶质在神经系统的发育以及突触可塑性和突触形成等过程中至关重要。胶质细胞在损伤后神经元的修复中发挥作用。在中枢神经系统(CNS)中,神经胶质抑制修复。称为星形胶质细胞的神经胶质细胞扩大并增殖形成疤痕,并产生抑制受损或切断的轴突再生的抑制性分子。在周围神经系统(PNS)中,称为施旺细胞的神经胶质细胞促进修复。轴突损伤后,施旺细胞恢复到较早的发育状态,以促进轴突的再生。 CNS和PNS之间的这种差异为CNS中神经组织的再生带来了希望。例如,在受伤或被遣散后,可能能够修复脊髓。施旺细胞也被称为神经白细胞。这些细胞通过将神经核内的神经纤维反复缠绕而包裹PNS的神经纤维。该过程产生了髓鞘,其不仅有助于导电,而且还有助于受损纤维的再生。
形成髓鞘
[编辑]在中枢神经系统中发现寡突胶质细胞,类似于章鱼:它们具有球状细胞体,最多具有15个臂状突起。 每个“手臂”伸出神经纤维并在其周围盘旋,形成髓鞘。 髓鞘可将神经纤维与细胞外液隔离,并加快沿神经纤维的信号传导。[18]
神经传导
[编辑]星形胶质细胞是三联突触的关键参与者。[19][20][21][22] 它们具有几个关键功能,包括从化学突触间清除神经递质,这有助于区分单独的动作电位,并防止某些神经递质(如谷氨酸)的毒性积累,否则会导致兴奋性中毒。 此外,星形胶质细胞响应刺激而释放神经胶质递质,例如谷氨酸,ATP和D-丝氨酸。[23]
临床意义
[编辑]历史
[编辑]胶质细胞是在1856年,由病理学家鲁道夫·魏尔啸,在他试图寻找脑中的“结缔组织”时所发现。
传统上认为人类脑中胶质细胞的含量大约为神经元的十倍。[17]在19世纪下叶胶质细胞被发现后,受到媒体报导的扭曲及断章取义,产生了“我们只使用了脑的十分之一”这个有名的迷思。胶质细胞借由管理突触间讯息的传递从而影响学习的角色,在最近才被发现(2004)。
其他图片
[编辑]-
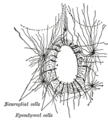 脊髓中央管的横切面,显示出室管膜细胞及其他胶质细胞。
脊髓中央管的横切面,显示出室管膜细胞及其他胶质细胞。 -
 一大脑叶(cerebellar folium)的横切面。
一大脑叶(cerebellar folium)的横切面。


注释
[编辑]- ^ Konstantinidis G. Elsevier's dictionary of medicine and biology: in English, Greek, German, Italian and Latin[M]. Elsevier, 2005. p1073.
- ^ 存档副本. [2023-02-17]. (原始内容存档于2023-02-17).
- ^ 3.0 3.1 Fields, R. Douglas; Araque, Alfonso; Johansen-Berg, Heidi; Lim, Soo-Siang; Lynch, Gary; Nave, Klaus-Armin; Nedergaard, Maiken; Perez, Ray; Sejnowski, Terrence; Wake, Hiroaki. Glial Biology in Learning and Cognition. The Neuroscientist. October 2014, 20 (5): 426–431 [2019-11-30]. ISSN 1073-8584. PMC 4161624
 . PMID 24122821. doi:10.1177/1073858413504465. (原始内容存档于2023-08-22).
. PMID 24122821. doi:10.1177/1073858413504465. (原始内容存档于2023-08-22).
- ^ Jessen KR, Mirsky R. Glial cells in the enteric nervous system contain glial fibrillary acidic protein. Nature. August 1980, 286 (5774): 736–7. PMID 6997753. doi:10.1038/286736a0.
- ^ 存档副本. [2023-02-17]. (原始内容存档于2023-08-06).
- ^ 存档副本. [2023-02-17]. (原始内容存档于2023-02-17).
- ^ 存档副本. [2023-02-17]. (原始内容存档于2023-02-17).
- ^ https://www.termonline.cn/word/1343722995876732929/1#s1
- ^ Wolosker H, Dumin E, Balan L, Foltyn VN. D-amino acids in the brain: D-serine in neurotransmission and neurodegeneration. The FEBS Journal. July 2008, 275 (14): 3514–26. PMID 18564180. doi:10.1111/j.1742-4658.2008.06515.x.
- ^ Swaminathan, Nikhil. Glia—the other brain cells. Discover. Jan–Feb 2011 [2019-11-30]. (原始内容存档于2014-02-08).
- ^ Gourine AV, Kasymov V, Marina N, et al. Astrocytes control breathing through pH-dependent release of ATP. Science. July 2010, 329 (5991): 571–5. PMC 3160742 . PMID 20647426. doi:10.1126/science.1190721.
- ^ Beltrán-Castillo S, Olivares MJ, Contreras RA, Zúñiga G, Llona I, von Bernhardi R, et al. D-serine released by astrocytes in brainstem regulates breathing response to CO2 levels.. Nat Commun. 2017, 8 (1): 838. PMC 5635109 . PMID 29018191. doi:10.1038/s41467-017-00960-3.
- ^ 13.0 13.1 von Bartheld, Christopher S.; Bahney, Jami; Herculano-Houzel, Suzana. The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting. The Journal of Comparative Neurology. 2016-12-15, 524 (18): 3865–3895. ISSN 1096-9861. PMC 5063692 . PMID 27187682. doi:10.1002/cne.24040.
- ^ Classic Papers. Network Glia. Max Delbrueck Center für Molekulare Medizin (MDC) Berlin-Buch. [14 November 2015]. (原始内容存档于2020-05-08).
- ^ Miyata, S; Furuya, K; Nakai, S; Bun, H; Kiyohara, T. Morphological plasticity and rearrangement of cytoskeletons in pituicytes cultured from adult rat neurohypophysis.. Neuroscience Research. April 1999, 33 (4): 299–306. PMID 10401983. doi:10.1016/s0168-0102(99)00021-8.
- ^ Rodríguez, EM; Blázquez, JL; Pastor, FE; Peláez, B; Peña, P; Peruzzo, B; Amat, P. Hypothalamic tanycytes: a key component of brain-endocrine interaction. (PDF). International Review of Cytology. 2005, 247: 89–164. PMID 16344112. doi:10.1016/s0074-7696(05)47003-5.
- ^ 17.0 17.1 Azevedo FA, Carvalho LR, Grinberg LT, et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of Comparative Neurology. April 2009, 513 (5): 532–41. PMID 19226510. doi:10.1002/cne.21974.
- ^ Saladin, Kenneth. Anatomy and Physiology, 6th Edition. McGraw Hill 2012. Page 446-448.
- ^ Newman, Eric A. New roles for astrocytes: Regulation of synaptic transmission. Trends in Neurosciences. 2003, 26 (10): 536–542. PMID 14522146. doi:10.1016/S0166-2236(03)00237-6.
- ^ Halassa MM, Fellin T, Haydon PG. The tripartite synapse: roles for gliotransmission in health and disease.. Trends Mol Med. 2007, 13 (2): 54–63. PMID 17207662. doi:10.1016/j.molmed.2006.12.005.
- ^ Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information.. Trends Neurosci. 2009, 32 (8): 421–31. PMID 19615761. doi:10.1016/j.tins.2009.05.001. hdl:10261/62092.
- ^ Santello M, Calì C, Bezzi P. Gliotransmission and the tripartite synapse.. Advances in Experimental Medicine and Biology 970. 2012: 307–31. ISBN 978-3-7091-0931-1. PMID 22351062. doi:10.1007/978-3-7091-0932-8_14.
- ^ Martineau M, Parpura V, Mothet JP. Cell-type specific mechanisms of D-serine uptake and release in the brain.. Front Synaptic Neurosci. 2014, 6: 12. PMC 4039169 . PMID 24910611. doi:10.3389/fnsyn.2014.00012.
参考文献
[编辑]- 陈子琏; 曾园山,张惠君. 人体结构学. 北京: 科学出版社. 2001. ISBN 7-03-009004-7.
- Brodal, Per. Glia. The central nervous system: structure and function. Oxford University Press. 2010: 19 [2016-05-18]. ISBN 978-0-19-538115-3. (原始内容存档于2013-06-18).
- Kettenmann and Ransom, Neuroglia, Oxford University Press, 2012, ISBN 978-0-19-979459-1 |http://ukcatalogue.oup.com/product/9780199794591.do#.UVcswaD3Ay4%7C (页面存档备份,存于互联网档案馆)
延伸阅读
[编辑]- Barres BA. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. November 2008, 60 (3): 430–40. PMID 18995817. doi:10.1016/j.neuron.2008.10.013.
- Role of glia in synapse development (页面存档备份,存于互联网档案馆)
- Overstreet LS. Quantal transmission: not just for neurons. Trends in Neurosciences. February 2005, 28 (2): 59–62. PMID 15667925. doi:10.1016/j.tins.2004.11.010.
- Peters A. A fourth type of neuroglial cell in the adult central nervous system. Journal of Neurocytology. May 2004, 33 (3): 345–57. PMID 15475689. doi:10.1023/B:NEUR.0000044195.64009.27.
- Volterra A, Steinhäuser C. Glial modulation of synaptic transmission in the hippocampus. Glia. August 2004, 47 (3): 249–57. PMID 15252814. doi:10.1002/glia.20080.
- Huang YH, Bergles DE. Glutamate transporters bring competition to the synapse. Current Opinion in Neurobiology. June 2004, 14 (3): 346–52. PMID 15194115. doi:10.1016/j.conb.2004.05.007.
- Artist ADSkyler(uses concepts of neuroscience and found inspiration from Glia)
外部链接
[编辑]- "The Other Brain" (页面存档备份,存于互联网档案馆)—The Leonard Lopate Show (WNYC) "Neuroscientist Douglas Field, explains how glia, which make up approximately 85 percent of the cells in the brain, work. In The Other Brain: From Dementia to Schizophrenia, How New Discoveries about the Brain Are Revolutionizing Medicine and Science, he explains recent discoveries in glia research and looks at what breakthroughs in brain science and medicine are likely to come."
- "Network Glia" (页面存档备份,存于互联网档案馆) A homepage devoted to glial cells.
- Role of glia in synapse development (页面存档备份,存于互联网档案馆)
- Overstreet L. Quantal transmission: not just for neurons.. Trends Neurosci. 2005, 28 (2): 59–62. PMID 15667925.
- Peters A. A fourth type of neuroglial cell in the adult central nervous system.. J Neurocytol. 2004, 33 (3): 345–57. PMID 15475689.
- Volterra A, Steinhäuser C. Glial modulation of synaptic transmission in the hippocampus.. Glia. 2004, 47 (3): 249–57. PMID 15252814.
- Huang Y, Bergles D. Glutamate transporters bring competition to the synapse.. Curr Opin Neurobiol. 2004, 14 (3): 346–52. PMID 15194115.
- New Source of Replacement Brain Cells Found (页面存档备份,存于互联网档案馆) - glial cells can transform into other cell types and reproduce indefinitely—tricks once thought exclusive to stem cells.
- Artist ADSkyler(uses concepts of neuroscience and found inspiration from Glia)
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||