
三疊紀
| 三疊紀 251.902–201.3百萬年前 | |

晚三疊世:2.2億年前的地球 | |
| 全時期平均大氣O 2含量 |
約16 Vol %[1] (為現代的80% ) |
| 全時期平均大氣CO 2含量 |
約1750 ppm[2] (為前工業時期6倍) |
| 全時期平均地表溫度 | 約17℃[3] (高於現代3℃)
|
直軸:百萬年前
三疊紀(英語:Triassic,符號T)是顯生宙中生代的一個紀,是中生代的第一個地質時代,位於二疊紀和侏羅紀之間,始於2.51億年前的二疊紀—三疊紀滅絕事件,終於2.01億年前的三疊紀—侏羅紀滅絕事件。雖然這段時間的岩石標誌非常明顯和清晰,其開始和結束的準確時間卻如同其它古遠的地質時代無法非常精確地被確定,其誤差在正負數百萬年。
三疊紀是四足動物演化歷史上的分水嶺時期,在這期間曾在古生代後期繁盛一時的離片椎類兩棲動物徹底衰落被邊緣化;合弓綱的獸齒類雖然作為劫後餘生的先鋒物種在三疊紀早期也曾繁盛一時,但經過一系列小規模滅絕事件後衰敗,僅存犬齒獸一支獨苗演化出了哺乳形類;蜥形綱的主龍類因其基於尿酸的泌尿系統更適應三疊紀日益乾旱的氣候,趁機迅速輻射崛起並演替搶占了陸地生態系統的上層生態位,使得蜥形綱首次壓倒合弓綱成為優勢物種;同時一些非主龍類爬行動物演化成為海洋爬行動物,作為首批重返水域生態系統的羊膜動物變成了海洋食物鏈中上層的掠食者,並掀起了對軟體動物演化影響深刻的中生代海洋革命。在晚三疊世早期的卡尼期洪積事件後,鳥頸類主龍崛起,開啟了恐龍和翼龍統治陸地長達1.4億年的時代。
詞源
[編輯]三疊紀的名稱是1834年弗里德里希·馮·阿爾伯提提出,他將在中歐普遍存在的位於白色的石灰岩和黑色的頁岩以及其間的三層紅色岩層統稱為「三疊」。今天,三疊紀被分成更多亞層。標誌三疊紀的典型的紅色沙岩說明當時的氣候比較溫暖乾燥,沒有任何冰川的跡象,今天一般認為當時在兩極沒有陸地或覆冰。因為當時地球上只有一個大陸,因此當時的海岸線比今天要短得多。三疊紀時遺留下來的近海沉積比較少,只有在西歐比較豐富,因此三疊紀的分層主要是依靠暗礁地帶的生物化石來分的。
分期
[編輯]三疊紀一般被分成早、中、晚三個世。國際地層委員會2018年8月版國際地層年代表將三疊紀劃分如下[6]:
| 三疊紀 (Triassic) | 晚三疊世 (Late Triassic) | 瑞替期 | 距今約208.5百萬年至距今201.3±0.2百萬年 |
| 諾利期 (Norian Age) | 距今約227百萬年至距今約208.5百萬年 | ||
| 卡尼期 (Carnian Age) | 距今約237百萬年至距今約227百萬年 | ||
| 中三疊世 (Middle Triassic) | 拉丁期 (Ladinian Age) | 距今約242百萬年至距今約237百萬年 | |
| 安尼期 (Anisian Age) | 距今247.2百萬年至距今約242百萬年 | ||
| 早三疊世 (Early Triassic) | 奧倫尼剋期 (Olenekian Age) | 距今251.2百萬年至距今247.2百萬年 | |
| 印度期 (Induan Age) | 距今251.902±0.024百萬年至距今251.2百萬年 |
古地理學
[編輯]
.png)
斯堪地那維亞半島
[編輯]古海洋學
[編輯]氣候
[編輯]三疊紀時的氣候普遍炎熱乾燥,這形成了現在可以看到的當時留下來的典型的紅色沙石。當時季節分明,有強烈的雨季。在兩極比較潮濕溫和。
植物群
[編輯]陸地植物
[編輯]浮游植物
[編輯]生物
[編輯]
三疊紀生物可以被分為三類:
海洋
[編輯]在海洋中新的珊瑚(例如:石珊瑚目)種類誕生,它們組成比較小的珊瑚群。頭足綱中的鸚鵡螺亞綱唯一倖存的一支恢復起來,形成新的種類(例如:鸚鵡螺科)。一開始的魚類種類比較少,說明只有少數種類於不久前的那次災難中倖存。海生爬行動物出現並繁盛。
陸地
[編輯]在大陸上繁盛的植物有蘇鐵、水韭、針葉樹和二叉羊齒。在北半球,針葉樹比較繁茂;在南半球,二叉羊齒是主要樹木。
- 喙頭龍目:一類雙孔類爬行動物,具有矮胖結實的身體與特別的喙狀嘴,具植食性。
- 植龍目:一類大型半水生掠食動物,具有長口鼻部與厚重鱗甲,在體型、外表、以及生活方式等方面與現代鱷魚類似。
- 堅蜥目:一類植食性偽鱷類演化支,身上有重骨板,體型由中到大,化石分布於三疊紀晚期地層。
- 勞氏鱷類:一類所知有限的偽鱷類集合群的非正式總稱,是三疊紀時期的陸地頂級掠食者
在二疊紀-三疊紀滅絕事件中倖存下來的生物開始繁衍生息。在三疊紀時期,主龍形下綱,尤其是主龍類逐漸取代了先前在二疊紀占主導地位的合弓綱。
偽鱷類的全盛期是晚三疊世,在這時代它們分為直立四肢的勞氏鱷科、植食性帶有裝甲的堅蜥目、大型狩獵動物波波龍科、小型敏捷的喙頭鱷亞目、還有其他一些小類群。
鳥頸類主龍是另一個重要的主龍演化支,在三疊紀時期分化為翼龍目和恐龍總目。然而恐龍在三疊紀並沒有占據陸地生態系統的優勢,占據陸地生態系統的動物是非鳥頸類主龍和屬於合弓綱的二齒獸類。







地理
[編輯]所有大陸都構成了一塊巨大的大陸:盤古大陸。在三疊紀期間,內陸水體的面積大大減少,除了海岸和河谷之外的大部分內陸地區具有沙漠景觀。盤古大陸從晚三疊世開始逐漸分裂[10]。 .
- 古地理復原
-
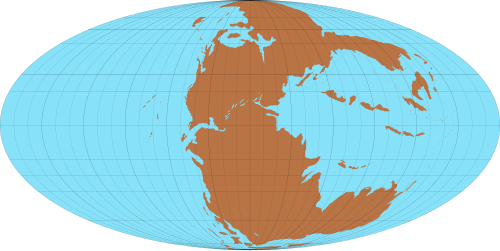 晚三疊世:2.2億年前
晚三疊世:2.2億年前
末期滅絕事件
[編輯]三疊紀以一次滅絕事件結束,尤其對海洋生物來說它的摧毀很慘重:牙形石滅絕,除魚龍和蛇頸龍外所有的海生爬行動物消失。腕足動物、腹足動物和貝殼等無脊椎動物受到巨大衝擊。在海洋中,22%的屬,大約一半的種消失。
這次滅絕事件並非在所有地方的摧殘程度都一樣。在有些地方幾乎沒有任何影響。在其它一些地方的大多數合弓類動物都消失了。許多早期的恐龍也均滅絕,而那些進步一些的恐龍卻倖存下來。許多主龍形下綱也都滅絕了。倖存的植物包括針葉類和蘇鐵等。
這次滅絕事件的原因還不清楚。在2.02至1.91億年前(利用氬40/氬39定年法[11]推定),盤古大陸開始分裂,導致了強烈的火山運動,這是地球大陸形成後最強烈的火山運動了。其它可能的原因有全球性的氣候冷卻或隕星。加拿大魁北克的一處隕石坑曾一度被認作是這次滅絕事件的起因,但後來的調查認為這個隕石坑是在2.14±0.01億年前形成的,隨後更精確的修正為2.0158±0.0028億年(兩者皆以鋯石中鈾衰變成鉛的比例來定年),比三疊紀的結束早了1000±200萬年,因此不太可能是這次滅絕事件的直接原因。[12]甚至連滅絕的確切時間也不十分確定。一些研究認為實際上當時有兩次滅絕事件,其相隔時間是1200至1700萬年。
這次滅絕事件為恐龍的發展提供了巨大的機會。恐龍在此後的1.4億年中成為了地球上最主要、種類最多和數量最大的陸地動物群。
參考文獻
[編輯]- ^ http://uahost.uantwerpen.be/funmorph/raoul/fylsyst/Berner2006.pdf
- ^ Image:Phanerozoic Carbon Dioxide.png
- ^ Image:All palaeotemps.png
- ^ McElwain, J. C.; Punyasena, S. W. Mass extinction events and the plant fossil record. Trends in Ecology & Evolution. 2007, 22 (10): 548–557. PMID 17919771. doi:10.1016/j.tree.2007.09.003.
- ^ Payne, J. L.; Lehrmann, D. J.; Wei, J.; Orchard, M. J.; Schrag, D. P.; Knoll, A. H. Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction. Science. 2004, 305 (5683): 506–9. PMID 15273391. doi:10.1126/science.1097023.
- ^ 國際地層委員會. 国际地层年代表 (PDF). Stratigraphy.org. [2018-09-01]. (原始內容存檔 (PDF)於2018-09-01).
- ^ Scheyer et al. (2014): Early Triassic Marine Biotic Recovery: The Predators' Perspective. PLoS ONE https://doi.org/10.1371/journal.pone.0088987 (頁面存檔備份,存於網際網路檔案館)
- ^ Prehistoric Life: The Definitive Visual History of Life On Earth. London: Dorling Kindersley. 2009: 206, 207. ISBN 9780756655730.
- ^ Douglas Palmer & Peter Barrett. Evolution: The Story of Life. London, Britain: The Natural History Museum. 2009. ISBN 9781845333393.
- ^ Н. В. Короновский, А. Ф. Якушова Основы геологии (頁面存檔備份,存於網際網路檔案館) 22.5. ПОДВИЖНЫЕ ПОЯСА (ПЕРЕХОДНЫЕ ЗОНЫ И ОКЕАНЫ
- ^ Nomade et al.,2007 Palaeogeography, Palaeoclimatology, Palaeoecology 244, 326-344.
- ^ Hodych & Dunning, 1992.

{kind=link}
{kind=link}