
海馬迴
| 海馬迴 | |
|---|---|
 | |
 海馬迴(圖下粉紅色部分)屬於邊緣系統 | |
| 基本資訊 | |
| 屬於 | 顳葉 |
| 標識字符 | |
| 拉丁文 | Hippocampus |
| MeSH | D006624 |
| NeuroNames | 3157 |
| NeuroLex ID | birnlex_721 |
| TA98 | A14.1.09.321 |
| TA2 | 5518 |
| FMA | FMA:275020 |
| 《神經解剖學術語》 [在維基數據上編輯] | |

海馬迴(英語:Hippocampus),是脊椎動物腦中的重要部分,其名字來源於這個部位的彎曲形狀貌似海馬。靈長類的海馬迴位於內側顳葉,擁有海馬角[1]及齒狀回等構造。哺乳動物都有兩個海馬迴,分別位於左右大腦半球。它是組成大腦邊緣系統的一部分,位於大腦皮質下方,擔當著關於短期記憶、長期記憶,以及空間定位的作用[2]。海馬迴中的位置細胞與內嗅皮層中的網格細胞在腦部進行空間定位和導航中扮演重要角色[3][4]。
在阿茲海默病中,海馬體是首先受到損傷的區域(海馬迴會腫大):表現症狀為記憶力衰退以及方向知覺的喪失。大腦缺氧(缺氧症)以及腦炎等也可導致海馬損傷。在動物解剖中,海馬體屬於腦的演化過程中最古老的一部分。來源於原皮質的海馬體在靈長類以及海洋生物中的鯨類中尤為明顯。雖然如此,與進化樹上相對年輕的大腦皮層相比,靈長類動物尤其是人類的海馬體在端腦中只占很小的比例。相對新皮質的發展,海馬體的增長在靈長類動物中的重要作用是使得其腦容量顯著增長。
研究史
[編輯]

解剖學家Giulio Cesare Aranzi(約1564年)首先使用海馬(hippocampus)一詞形容這一大腦器官,源於此部位貌似海馬。這一部位最初被認為司控嗅覺,而非現在周知的記憶儲存作用。俄國學者Vladimir Bekhterev於1900年左右基於對一位有嚴重記憶紊亂的病患者的長期觀察,首先提出海馬體與記憶相關。但是,其後的很長時間,學界習慣上關於海馬體的作用都被認為和其他大腦邊緣系統一樣,司控情緒。
1957年Scoville與Milner關於著名的病人H.M.的病例報告引起了眾多科學家的關注,並使人開始認識到海馬體對記憶起重要作用。為減輕H.M.時常發作的癲癇症狀,其腦內側顳葉被切除(包括當中的兩個海馬迴),由此導致了一系列的相關空間以及時間的記憶損傷。重要的是HM仍然能完成程式性任務的學習(這一點與紋狀體相關聯),甚至有著高於常人水平的智商。H.M.的智能與陳述性記憶展現出顯著的分裂。絕大多數的哺乳類動物海馬體的大小取決於腦容量的大小,但是鯨類這一部位的發育相對不完全。
研究成果
[編輯]海馬體是哺乳類動物的中樞神經系統中的腦的部分(大腦皮質)中被最為詳細研究過的一個部位。在解剖學以及組織學上,海馬具有一目了然的明確構造。海馬內部有形成形態美觀的層面。也就是神經細胞的細胞體與其神經網區域呈層狀排列。
20世紀初,開始有科學家發現海馬對於某些記憶以及學習有著基本性的作用。特別是1957年Scoville和Milner報告了神經心理學中很重要的一個病例。這是來自一位被稱為H.M.的病人(亨利·古斯塔·莫萊森)的病者報告,亨利·莫萊森算是神經學領域中被檢查得最詳細的人。由於亨利長期被癲癇症狀困擾,神經外科醫生斯科維爾 (William Scoville)決定為他進行手術,他在他額頭兩側做了局部麻醉,鑽了兩個小孔,用一根金屬吸管吸出大部分海馬組織(Hippocampus)以及海馬周圍的部分內側顳葉組織。手術後,亨利的癲癇症狀被有效控制,但自此以後失去了形成新的陳述性長時記憶的能力。這個發現令許多人想進一步了解海馬區在記憶及學習機制的作用,因而成為一種流行,無論在神經解剖學、生理學、行為學等等各種不同領域,都對海馬區做了相當豐富的研究。現在,海馬區與記憶的關係已經為人所知。
許多人對海馬區與癲癇發作的關係也有很濃厚的興趣。海馬區在腦中為發作閾值低的部位。因為幾乎所有癲癇患者的發作皆由海馬區所起始,像這類以海馬區為主的發作,有許多時候是很難以藥物治療的。而且,海馬區中有一部分,尤其是內嗅皮質,為阿爾茲海默氏症最先產生病變的地方,海馬區也顯示出容易因貧血、缺氧狀態而受損傷。
海馬區在解剖學以及機能構造上都是其它大腦皮質系統的研究樣本。大腦皮質在最近開始被關注與研究,現在已知的關於中樞神經系統的突觸傳導的見解多受益於海馬區的研究。而海馬區的相關知識則多源於齒狀回與海馬的標本。
解剖結構
[編輯]海馬迴,是被稱作「海馬區」(hippocampal region)的大腦邊緣系統的一部分。海馬區可分為:齒狀回(dentate gyrus)、海馬迴、下托(subiculum)、前下托(presubiculum)、傍下托(parasubiculum)、內嗅皮質(entorhinal cortex)[5]。這之中齒狀回、海馬迴、下托的細胞層為單層,合稱「海馬結構(hippocampal formation)」,其上下夾有低細胞密度層和無細胞層。此外的部位有複數的層面構成。齒狀回與海馬的單層構造對神經解剖學以及電生理學的研究進步作出了貢獻。

儘管關於海馬與其向鄰近的大腦皮層的表述尚缺乏一致的觀點,通常情況下術語上的「海馬結構」指的是齒狀迴,CA1-CA3部位(或CA4,常稱為hilus區並被認為是齒狀迴的一部分),以及腦下腳(另見阿蒙神之角)。CA1與CA3部位構成嚴格意義上的海馬。
資訊進入海馬時由齒狀回流入CA3到CA1再到腦下腳,在每個區域輸入附加資訊在最後的兩個區域輸出。CA2隻占海馬的一個很小部分通常將其對海馬的功能忽略,值得注意的一點是這一小區域似乎能抵抗由於例如癲癇等造成的大規模的細胞破壞。
人們普遍認為這些區域的每一部分在海馬的資訊處理過程中都扮演著一個具有獨特功能的角色,但迄今為止對每一區域功能的細節還不甚了解。
一般記憶中的作用
[編輯]
心理學家與神經學家對海馬的作用存在爭論,但是都普遍認同海馬的重要作用是將經歷的事件形成新的記憶(情景記憶或自傳性記憶)。一些研究學者認為應該將海馬看作對一般的陳述性記憶起作用內側顳葉記憶系統的一部分(陳述性記憶指的是那些可以被明確的描述的記憶,如「昨天晚飯吃了什麼」這樣的關於經歷過的事情的情景記憶,以及「地球是圓的」這樣的關於知識的概念記憶)。
有跡象顯示,雖然這些形式的記憶通常能終身持續,在一系列的記憶強化以後海馬便中止對記憶的保持。海馬的損傷通常造成難以組織新的記憶(順行性失憶症),而且造成難以搜尋過去的記憶(逆行性失憶症)。儘管這樣的逆行性效果通常在腦損傷的很多年之前就開始擴展,一些情況下,相對久遠的部分記憶能夠維持下來。這表明海馬將鞏固以後的記憶轉入了腦的其他的部位。但是,舊的記憶是如何儲存的要用實驗來檢測的話存在一些難點。另外,在一些逆行性失憶症案例中,在海馬遭受損傷的數十年前的記憶也受到了影響,導致了這一關於舊的記憶的觀點的爭議。
海馬的損傷不會影響某一些記憶,例如學習新的技能的能力(如學習一種樂器),將設這樣的能力依靠的是另外一種記憶(程式記憶)和不同的腦區域。有跡象表明著名的病人HM(作為治療癲癇病的手段他的內側顳葉被切除)有組織新的概念記憶的能力。
空間記憶以及方向定位中的作用
[編輯]

有些證據提供以下的線索:空間訊息的儲存與處理牽涉到海馬迴。在老鼠的海馬區里,插一根記錄單個神經細胞的電極(electrode),讓老鼠在一個開放式的試驗區域自由的跑動,老鼠沿高架軌道來回跑動,停在各端吃小食品作為獎勵,同時記錄神經細胞的動作電位 (action potential)和老鼠跑的路線及位置,當老鼠跑到實驗房間的某個地方的時候,海馬迴內的某一個特定的神經細胞A就會動作 (active),而周圍的其他細胞是不動作 (inactive)的,而跑到其他地方的時候,這個A細胞就不會動作 (inactive),而另外一個或多個細胞就會放電 (fire),這種對自身在環境中所處位置而產生對應反應的神經細胞,被稱為定位細胞(place cells)。對於每個 place cell 所對應的實際位置區域便被定義為 place field。通過一系列的實驗約翰·奧基夫 (O』Keefe) 推測,這些 space cells 組合在一起,形成了一個認知地圖(cognitive map)。通過和其他在海馬迴內以及周圍區域的神經細胞一起合作,place cells 其著如同一張地圖的作用。老鼠實驗的研究顯示,海馬迴的神經元有空間放電區,這些細胞稱為地點細胞(place cells)。如果老鼠發現自己處在某個地點,不論該老鼠移動的方向為何,有些細胞會發電,而大部分的細胞至少會對頭的方向、移動方向感到敏感。在老鼠身上,有些細胞稱為分野細胞(splitter cells),該種細胞的發電取決於動物的近期經驗(回顧記憶;retrospective memory)、或是期待即將的為來(前瞻記憶;prospective memory)。根據不同的身處地點,不同的細胞會發電;因此,只要觀察細胞的發電情形,就可能指出動物身處的地點。在人類身上,當人們在虛擬世界的城鎮裡在尋找方向時,就會牽涉到「地點細胞」。這樣的發現是源於如下的研究:在嚴重癲癇患者的大腦裡面植入電極,當作是患者在手術過程中診斷的方式。
發現了「地點細胞」,讓世人覺得海馬迴可能扮演「認知地圖」(cognitive map)的角色,而認知地圖就是環境格局的神經重現。然而,針對這樣的觀點,近期的證據提出懷疑,並且指出海馬迴對於「尋找方向」(navigation[來源請求])更根本的過程非常重要。儘管如此,動物實驗顯示,即使要完成簡單的空間記憶活動,健全的海馬迴是必要的(譬如把目的地藏住,要動物找路回去)。
若海馬迴不健全,人類可能就無法記住曾經去過的地方、以及如何前往想去的地點。研究人員相信,若要在熟悉環境之間找出捷徑、以及新的路線,海馬迴扮演極重要的角色。針對這樣尋找方向的能力,有些人比其他人能力強;此外,大腦顯影研究顯示,這些尋找方向能力比較好的人,在尋找方向時,他們的海馬迴比較活躍。

另外,倫敦計程車司機必須要記住很多地點,並且知道這些地點之間最直接的路線(他們必須通過嚴格的考試,該考試名為「知識」,英文名是「The Knowledge」,才能得到倫敦著名的黑色計程車「black cab」的駕駛執照)。在倫敦大學學院[6]的研究顯示,相較於一般民眾,倫敦計程車司機的海馬迴體積較大,至於更有經驗的計程車司機的海馬迴體積又更大。然而,有較大的海馬迴是否有助於成為計程車司機、或是成為計程車司機或以找捷徑為生是否能夠使得一個人的海馬迴變大仍待研究。
在印第安那大學進行的老鼠實驗提出了如下的可能性:在反覆的迷宮實驗裡觀察老鼠的表現,海馬迴的型態跟「兩性異形」息息相關。對於將地點空間化、找出自己所在,公老鼠表現比較好,因為公老鼠的海馬迴體積比較大。
腦中定位系統
[編輯]2014年諾貝爾生理學或醫學獎授予了約翰·奧基夫 (John O'Keefe)和莫澤夫婦、邁-布里特·莫澤 (May-Britt Moser)、 愛德華·莫澤 (Edvard Moser)。 奧基夫在1971年發現了海馬區中的位置細胞 ,莫澤夫婦在2005年發現了內嗅皮層中的網格細胞 ,它們構成了腦中定位系統的細胞基礎(cells that constitute a positioning system in the brain)。[3]
圖片
[編輯]-
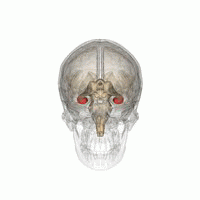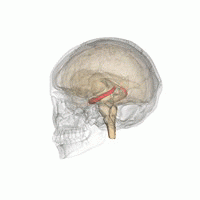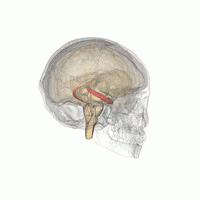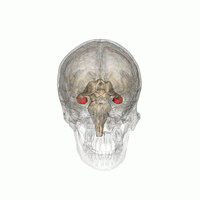 從不同的角度觀看紅色海馬的位置。
從不同的角度觀看紅色海馬的位置。 -
 海馬在腦的部位。
海馬在腦的部位。 -
 海馬的基本路線,DG:海馬齒狀回、Sub:海馬下托區、EC:內嗅皮質。
海馬的基本路線,DG:海馬齒狀回、Sub:海馬下托區、EC:內嗅皮質。 -
 尼氏染色獼猴大腦的冠狀切面,(圓圈)顯示海馬。
尼氏染色獼猴大腦的冠狀切面,(圓圈)顯示海馬。 -
 大腦的海馬區(左)和海馬(右)。
大腦的海馬區(左)和海馬(右)。

.png)


參考資料
[編輯]- ^ Pearce, 2001
- ^ Wright, Anthony. Chapter 5: Limbic System: Hippocampus (頁面存檔備份,存於網際網路檔案館). Department of Neurobiology and Anatomy, The UT Medical School at Houston
- ^ 3.0 3.1 The Nobel Prize in Physiology or Medicine 2014. NobelPrize.org. [2021-06-20]. (原始內容存檔於2018-08-10) (美國英語).
- ^ Lisman, John; Buzsáki, György; Eichenbaum, Howard; Nadel, Lynn; Ranganath, Charan; Redish, A. David. Viewpoints: how the hippocampus contributes to memory, navigation and cognition. Nature Neuroscience. 2017-11, 20 (11): 1434–1447. ISSN 1546-1726. doi:10.1038/nn.4661 (英語).
- ^ Schultz, Christian; Engelhardt, Maren. Anatomy of the hippocampal formation. Frontiers of Neurology and Neuroscience. 2014, 34: 6–17 [2021-06-26]. ISSN 1662-2804. PMID 24777126. doi:10.1159/000360925. (原始內容存檔於2021-07-02).
- ^ Macguire et al, 2000
- Brain Systems Underlying Declarative and Procedural Memories in Neuroscience second edition by Dale Purves, et al (2001) Published by Sinauer Associates, Inc. ISBN 0-87893-742-0
- Mizuno K. and Giese K.P. Hippocampus-dependent memory formation: do memory type-specific mechanisms exist?. Journal of Pharmacological Sciences. 2005, 98 (3): 191–7. doi:10.1254/jphs.CRJ05005X.
- Broadbent N.J., Squire L.R. and Clark R.E. Spatial memory, recognition memory, and the hippocampus. Proceedings of the National Academy of Sciences of the United States of America. 2004, 101 (40): 14515–20. doi:10.1073/pnas.0406344101.
- Macguire, E.A.; et al. Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences of the United States of America. 2000, 97 (8): 4398–4403. doi:10.1073/pnas.070039597.
外部連結
[編輯]{kind=link}
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||