
烏鵰
| 乌雕 | |
|---|---|

| |
| 印度 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 鸟纲 Aves |
| 目: | 鹰形目 Accipitriformes |
| 科: | 鹰科 Accipitridae |
| 属: | 花鵰屬 Clanga |
| 种: | 乌雕 C. clanga
|
| 二名法 | |
| Clanga clanga | |

| |
| Range of C. clanga
繁殖區 過境 非繁殖區
| |
| 異名 | |
|
Aquila clanga | |
乌雕(学名:Clanga clanga)为鹰科花鵰屬的鸟类,又名花雕,是一種大型遷徙猛禽,屬於鷹科。
牠是真鵰亞科的一員[3]。曾經被歸類於鵰屬(Aquila ),但現在已與另外兩種花鵰一起重新歸類到不同的花鵰屬屬[4]。
在繁殖季節,花雕廣泛分佈在東歐、中歐部分地區、俄羅斯中部、中亞和中國部分地區以及其他偏遠地區。在冬季,牠們會遷徙,主要遷往南亞、東南亞、中東、地中海盆地北部以及東非部分地區[1][5]。花鵰比大多數其他靴鵰更喜歡潮濕的棲地,偏好河岸帶以及沼澤、湖泊、池塘和其他被林地包圍的水體。牠們主要在有高水位的洪氾區繁殖。在冬季和遷徙期間,牠們經常尋找類似的濕地棲地,但也有在乾燥的高地如稀樹草原高原上被觀察到[5][6]。
花鵰是機會主義的覓食者,尤其在冬季。牠會很樂意拾食各種食物來源,包括腐肉,以及小型哺乳動物(主要是囓齒動物)、青蛙和各種較小的鳥類(特別是水鳥),有時也捕食爬行動物和昆蟲。花鵰主要是空中獵食者,從隱蔽的棲木滑行到沼澤或濕地捕獵獵物[5][6][7][需要解释]。
該物種在大樹上築巢,巢由樹枝構成,通常產下一至三顆蛋。雌鳥負責孵卵和育雛,雄鳥負責狩獵並提供食物。父母每年很少撫養超過一隻幼鳥[8][9]。如同少數猛禽物種,最年長的雛鳥通常比其他雛鳥大得多,且常常會攻擊並殺死較小的兄弟姐妹[10]。
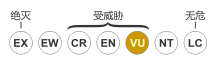
花鵰的分佈範圍與密切相關的小斑鵰 (Clanga pomarina) 有重疊。這兩個物種經常交配並產生混種後代,這對於數量稀少的花鵰族群有害[11]。花鵰被國際自然保護聯盟列為易危物種[1]。牠們的族群面臨棲地破壞、與物體碰撞以及與小花鵰雜交的威脅[5][12]。
分類學與詞源
[编辑]
花鵰是真鵰亞科(或「靴鵰」)的成員,這是一個單系群,屬於較大的鷹科家族。所有靴鵰的腿部都覆有羽毛。這個多樣化且分佈廣泛的家族成員可在除南極洲外的所有大陸上找到。靴鵰已確認有38個物種[3][5][13]。
靴鵰通常與鵟屬(Buteo)和漁雕屬(Haliaeetus)及其他體型較重的鷹科成員歸為一組,但牠們可能與體型較修長的鷹形目鷹亞科比以前認為的更為接近。花鵰最接近的現存親緣物種是小斑鵰。據信牠們在約370萬年前的中新世末期從其最近共同祖先分化而來。這種「原始花鵰」可能生活在現代阿富汗地區,並在最後冰河期開始時,因為中亞地區冰川和沙漠的進退而分化為南北兩支。北支後來分化為今天的花鵰(東部)和小斑鵰(西部),這一過程可能發生在中新世晚期與更新世邊界,接近200萬年前[14][15][16]。
花鵰曾被歸類為鵰屬(Aquila)的成員,與幾種大多數為棕色的大型鵰屬於同一類[5]。然而,利用一個線粒體和兩個核基因序列進行的分子系統發生研究顯示,花鵰組成了一個單系群,與高冠鷹鵰 (Lophaetus occipitalis) 並列。研究顯示,花鵰應該歸類為高冠鷹鵰屬 (Lophaetus),或者將這些物種全部歸類為鵰屬(Aquila)[3][4][17]。此外,發現花鵰與亞洲本土的林鵰 (Ictinaetus malaiensis) 之間有密切關係。花鵰、高冠鷹鵰和林鵰可能構成一個複合種或支系[18]。由於壓倒性的基因證據以及花鵰與其姊妹分類群在形態和生態上的巨大差異,花鵰最終被重新歸類為一個獨立的花鵰屬(Clanga)[18][19]。其學名Clanga可能來源於古希臘語 κλαγγή(直译:「叫聲」),[20]或其根源可能是希臘語單詞klangos(plangos的變體形式),意為亞里士多德提到的一種「鷹」[6]。
由於這兩個物種的親緣關係在所有經過密切研究的鷹科(Accipitrid)分類群中可能是最接近的,儘管有顯著的基因差異,但花鵰與小斑鵰之間的廣泛雜交現象仍然存在[18][19][21]。這兩個物種的線粒體基因序列差異超過3%,這幾乎是區分兩個物種的最低基因差異的兩倍[22]。第三個花鵰物種印度花鵰 (Clanga hastata),於2006年被確認為與大小花鵰相似的獨立物種[16][23]。
描述
[编辑].jpg)

花鵰體型相當大且結實。通常為黑褐色,並具有明顯對比的黃色喙。該物種擁有短頸部、大且經常毛茸茸的後頸、強壯的喙以及短的嘴裂線,鼻孔呈圓形。翅膀寬闊且長,達到尾端。尾巴相對較短且呈圓形[5][8]。寬闊的翅膀和短尾巴的整體效果使牠們的輪廓幾乎像禿鷲一樣。牠們的腳掌較大,覆蓋腿部的羽毛比小花鵰的排列得較鬆散[5][6]。
花鵰通常會停棲在開闊處。大多數棲木位於森林邊緣的樹頂,或者是更為孤立的觀察點,如灌木、電線杆或陡峭的河岸。花鵰經常在地面上覓食,或以稍微駝背的姿勢在地面上休息。
成年花鵰通常呈相當一致的深棕色至黑褐色,儘管顏色可能顯得帶有紫色和光澤。牠們在剛換羽時可能顯得更加鮮明對比[5][6]。
上翼的覆羽通常比其他部分顏色稍淡,雖然這些鷲通常顯得色調一致,但有兩個對比鮮明的特徵:淺色的喙和尾巴上方的狹窄白色U形標記,儘管後者通常在靜止時隱藏不見。該物種具有兩性異形,甚至可能具有生物多態性。淺色和中間型表現型比較罕見,但在東部地區稍微更為常見[5][24]。
有時被稱為Clanga clanga fulvescens的淺色成年體具有雙色羽毛。尾部、飛羽和大覆羽均為黑色,而身體及其餘的覆羽呈現淺黃色或淺金黃色,有時隨著年齡增長變為奶油色。fulvescens表現型的淺色通常與眼睛周圍、翅膀前緣以及胸部較少且稀疏的暗色形成對比[5][25]。中間型和其他變種非常罕見,但包括身體略淡、上前翼覆羽上具有可變的黃棕色條紋或斑駁(使牠們看起來像小花鵰的幼鳥),或具有深色條紋胸部和淡尖覆羽的黃褐色斑駁體(像白肩鵰 (Aquila heliaca)的幼鳥)。這些中間型可能在上身顯示典型的深褐色至黑色,但在飛行中顯示出淺色斑駁的灰色翼襯,或除了對比鮮明的淺色腹部外,其他部分呈現正常顏色[5][6][25]。
幼年花鵰通常是均勻的黑褐色,並有白色至黃色的滴狀斑點。一些幼鳥全身布滿斑點,而另一些則較少。牠們一般顯示出沿上翼覆羽的明顯斑點,形成明顯的翼條和尾羽,只有外初級飛羽例外。幼鳥羽毛下方通常有寬廣的奶油色尖端,尤其是在兩側和腿部下方經常顯示出一些淡褐色條紋。fulvescens型花鵰的幼鳥類似於淺色成年體,但顯示出典型幼鳥的重點斑點翅膀和尾巴,並經常在肩胛羽和中覆羽上顯示一些較暗的中心。到第二至第三年,羽毛通常已經相當磨損,但白色的尖端仍然形成足夠顯著的翼條(與小花鵰不同),直到第二個冬季,大多數覆羽會新換羽並有較小的淺色尖端。從大約第三年中期開始,羽毛更像成鳥,僅留下少量或無不明顯的斑點,但飛羽年齡不一,看起來凌亂[5][25]。亞成體通常顏色更為一致,但通常仍會顯示出一些大覆羽的淺色尖端[5]。大約在第五年左右達到成熟,但有時直到第六年才開始繁殖[5][26]。裸露部位在不同年齡下顏色變化不大,眼睛為深棕色,而喙和腳在所有年齡下都為黃色。
在飛行中,這是一隻體型大、顏色深的猛禽(通常看起來比實際尺寸更大),具有明顯突出的頭部、長翅膀(由於翅膀寬廣,常常看起來較短)、略微突出的次級飛羽,以及相對方正的七指翼尖,儘管幼鳥的翅膀可能看起來更圓[5]。在飛行時,花鵰顯得身軀沉重,經常懸掛在翅膀下方,且擁有相對短而寬的尾巴。牠們的翅膀拍動快速,幾乎沒有向上揮動的動作,飛行動作比亞洲草原鵰(Aquila nipalensis)輕盈,但相對於小斑鵰,顯得稍微笨重、優雅度較低,也不像鵟屬(Buteo)那樣[5]。花鵰在翱翔時,翅膀幾乎是平的,翼尖常常微微向下,且初級飛羽充分展開。在滑翔時,翅膀略微彎曲,手臂和翼尖之間形成明顯角度,強調了翅膀的短小外觀[5]。
在上翼,花鵰的主要羽軸由白色基部構成,並部分呈淺色外網,形成了淺色斑塊,這在各年齡段都可見到,但成鳥的斑塊較小且不太明顯[5]。下翼幾乎一般有一條由最外側三根初級飛羽、次級飛羽和最內側初級飛羽的白色基部形成的單一白色新月形斑紋。近距離觀察時,可以看到九到十一條密集且狹窄的暗色條紋逐漸向翼尖處消失[5]。飛行時,成鳥通常為均勻的黑色,有微弱的淺色U形標記位於尾部上方,翼覆羽略淺,羽軸較淺。成鳥有時會有略淺的翼內襯,類似於小斑鵰,但在初級飛羽基部只有單一的(而不是雙重的)白色新月形斑紋。fulvescens(淺色)型成鳥的翼覆羽和身體在兩側面上對比鮮明地呈現為淡黃褐色至黃褐色[5][27]。
幼鳥在飛行時通常顯得非常暗色,上下均有大量斑點,但有些幼鳥的斑點僅限於翅膀、肩胛羽和腿部。所有幼鳥在良好觀察下都會顯示出翼覆羽上特有的白色末端斑點,形成兩到三條翼條[5]。此外,幼鳥的翅膀和尾巴具有奶油色的後緣。下方,幼年花鵰的翼襯大部分為黑色(除了奶油色的下尾覆羽),與較淺的灰色煙羽形成對比。其他幼鳥的羽色較淡,但羽軸與典型幼鳥的相似[5]。
在伏爾加河–烏拉爾山脈地區繁殖的鳥類體型略大,羽毛特徵更為低調。個體略小的鳥類似乎在歐洲更西部地區較為普遍。冬季在印度的鳥類比中東地區的鳥類體型大約有5%的差異[28]。
體型
[编辑].jpg)
花鵰屬於中等大小的鷹,但也是一種大型猛禽[8]。該物種顯示出明顯的兩性異形,雌性在體型上較大,儘管兩性羽毛相似。體型差異線性可達26%,雌性體重可達雄性兩倍之多,這使得牠們可能與冠鷹(Polemaetus bellicosus)成為真鵰亞科(Aquilinae)中兩性異形最明顯的成員[5][8]。然而,最大的雄性花雕在大多數線性和質量測量中可能與較小的雌性重疊。[25][29]。完全成長的花鵰總長度可變化59至74 cm(23至29英寸)[30][31]。翼展方面,雄性據報測量為155—177 cm(61—70英寸),而雌性可測量167—185 cm(66—73英寸)[8][30][32][33]。雄性體重據報範圍為1,537至2,000 g(3.389至4.409磅)[8][29][34][35],雌性體重範圍為1,820至3,250 g(4.01至7.17磅)[8][29][35]。在標準測量中,翼弦雄性範圍為477—519 mm(18.8—20.4英寸),雌性為507—545 mm(20.0—21.5英寸)[8][5][34][36]。短尾長度在雄性範圍為227—249 mm(8.9—9.8英寸),雌性為235—268 mm(9.3—10.6英寸)[5]。相對較長的脛跗骨在雄性範圍為97—105 mm(3.8—4.1英寸),雌性為96—112 mm(3.8—4.4英寸)[5][37]。據報導,喙峰長度可變化32.5至39 mm(1.28至1.54英寸)[38]。
鳴聲聲
[编辑]花鵰在繁殖時相當吵鬧,且在冬季時尤其是在小群體中常常非常聒噪[5]。最常聽到的叫聲是在同種衝突期間發出的柔和、單音節、穿透力強、高音且急促的哨聲,各種記錄為kyack、kluh、tyuck或dyip。這種叫聲與小斑鵰的叫聲類似,但音調稍微低沉且更響亮。這些叫聲比亞洲草原鵰的叫聲高亢,並且比白肩鵰的叫聲(Aquila heliaca)高得多,後者的叫聲帶有喉音,讓人聯想到青蛙的叫聲[5][8]。此外,類似的三音節吠聲似乎用來警告餵食地點的入侵者,有時被認為是一種刺耳的chrr-chrr-chaa-chaa、kyak-yak-yak和kyew-kyew-kyew。反覆的叫聲累積起來的效果被比作「小獵犬」的叫聲[8][34][39]。與許多猛禽一樣,雌鳥的音調較低沉且更沙啞[8]。有一隻花鵰在兩天的錄音中發出了一種異常響亮的叫聲,聽起來與大冠鷲(Spilornis cheela)的典型叫聲的前兩個音節非常相似[6]。
識別
[编辑]
辨認花鵰在野外可能相當困難[40]。該物種主要根據其結構和比例來區分於小斑鵰,儘管遠距離觀察時可能幾乎無法分辨[5][24]。與小斑鵰相比,成鳥顯得翅膀非常寬,這使得頭部顯得相對較小。然而,花鵰的幼鳥看起來可能不那麼笨重,翅膀較窄,翅膀輪廓更圓,尾巴較長,使其比例更接近小斑鵰[5][24]。
花鵰可以明顯地在體型上更大,雌鳥尤其明顯地比大多數小斑鵰大得多,但這兩個物種之間有很大的體型重疊。在某些情況下,雄性花鵰幾乎不比雄性小斑鵰大[5][6]。並排比較時,花鵰通常顯著比小斑鵰顏色更深[30][24],且其尾上覆羽較深色,沒有頸背斑,黑褐色的均勻雙翼和上翼覆羽(而非對比鮮明的鐵鏽色棕褐色)也是顯著特徵[24]。儘管困難,但可根據形態、通常較暗的翼內襯、初級羽斑和腕弧的外觀差異來區分中間型花鵰與年幼的小斑鵰[5]。幼鳥通常缺少小斑鵰的淺色頸背斑,但有時會出現,「儘管僅比其餘羽毛稍淡,且從不呈現赭色或橙色」[41]。典型地,幼年花鵰的斑點和橫斑圖案更強,但這並非可靠的區分[24]。
花鵰與小斑鵰的雜交種更難區分,且其外觀常常混淆不清,有些雜交種在外觀上更接近其中一個物種[24]。可通過手持測量如喙高、寬以及幼鳥白斑的範圍和中趾長度來區分純花鵰和純小斑鵰[42][43]。
在印度次大陸,花鵰可能與印度花鵰混淆[6]。印度花鵰體型較小(類似於小斑鵰),翅膀較窄且尾巴較長,初級飛羽指間更深切且方形末端。印度花鵰的初級飛羽窗較明顯,腹部顏色較淺且斑紋不明顯,上身較淺(顏色更像草原雕),大翼覆羽的淺色末端較不明顯且分散。此外,印度花鵰的嘴裂明顯更深,延伸至眼後[6][30]。
與類似或更大尺寸的非花鵰相比,花鵰通常在特徵上較為緊湊,具有相對寬(且看起來較短)的翅膀、較短的尾巴,以及整體較暗且圖案分明的羽毛[6]。亞洲草原雕與花鵰相似,但體型更大且更笨重,而花鵰的頸部較短,喙較小且嘴裂線較短,無頸背斑(見於成年的亞洲草原雕),褲腳較窄且不鬆散,翅膀普遍較短且略寬。儘管在遠距離看來類似,亞洲草原雕在灰色飛羽上有更明顯、更廣泛的橫斑,完全缺乏腕弧,下喉和頸背較淺,初級羽斑較大但更模糊[5][30]。
fulvescens 和中間型的花鵰類似於許多老鷹,但可通過其翼下顏色和圖案(如其明顯的腕弧和暗色、細緻的羽軸斑紋)來區分,這與類似體型的草原鵰(Aquila rapax)的淺色或中間型變異不同,後者通常背色較淺且無散亂的暗色面孔,且具有典型的較窄翼比例。白肩鵰的幼鳥可能與fulvescens花鵰相似,但體型更大且結構上不同。白肩鵰的翅膀更長且較窄,頸部更長,喙更大且突出,喙甲呈長方形(而非橢圓形),嘴裂線較長且狹窄,明顯的淺色內初級飛羽,無腕弧,胸部有褐色條紋(儘管花鵰可能顯示一些散亂的標記),腿部羽毛無標記,眼睛為淺色,內初級飛羽上有明顯的淺色窗[5][44]。除了結構上的不同,亞成年亞洲草原雕可通過其較厚的羽軸斑紋和較淺的下翼對角線來區分於較淺型的花鵰[5]。在分佈範圍的東部,可以將其與更暗的林鵰區分開來,後者體型較纖細,具有槳狀翅膀和長且明顯橫斑的尾巴[5]。
分佈與棲地
[编辑]
繁殖範圍
[编辑]這種猛禽主要繁殖於古北界和東洋界地區[5]。分布于北欧、欧洲东南部和亚洲大陸。分布于台灣本島以及中国大陆的新疆、内蒙古、东北、长江下游、河北、河南、浙江、福建、广东、广西等地,主要栖息于连接水源的森林草原。该物种的模式产地在西伯利亚。[2]
它也在中部和東南歐繁殖,但僅限於波蘭、白俄羅斯、烏克蘭、保加利亞[45]、羅馬尼亞、塞爾維亞和匈牙利的小型、非連續分布區域[1][21][46]。在東歐開始有較連續的繁殖範圍,涵蓋愛沙尼亞東部、拉脫維亞和立陶宛東部,以及白俄羅斯中部[1]。部分繁殖鳥隻不確定是否會擴展至芬蘭東部邊緣[47]。在俄羅斯歐洲部分廣泛分布,適合的棲地從阿爾漢格爾斯克州的大部分地區延伸到白海的下部海岸。它們遍佈中俄羅斯的大部分地區,北部的可能邊界在舒雷什卡爾斯基區和皮特基亞蘭塔區[1][34][48]。其在俄羅斯以外的分佈範圍包括哈薩克斯坦北部的大部分地區,並在東哈薩克斯坦州和哈薩克斯坦南部有已知的孤立繁殖區。花鵰還在從吉爾吉斯斯坦和毗鄰的俄羅斯地區延伸至中國新疆的孤立區域繁殖[1][48][49]。有時候,花鵰也會在印度次大陸繁殖,據報從古吉拉特邦向北到旁遮普省,其繁殖範圍記錄從南至索拉什特拉地區,北至馬哈拉施特拉邦。然而,這可能僅僅是歷史記錄,今天幾乎可以肯定不存在穩定的繁殖種群[6][50][51][52]。它們也在蒙古北部繁殖,並深入到中國東北和北韓北部[1]。
遷徙範圍
[编辑]花鵰在遷徙期間廣泛分布,通常秋季在九月至十一月間,春季在二月至四月間。遷徙期間的分布範圍比繁殖或越冬期間更為廣泛[1][6][21]。遷徙時,花鵰可能出現在東歐的大部分地區、安納托利亞和整個中東、中亞(從哈薩克斯坦南部開始)及南亞西部[1][21][53][54]。
這個物種容易出現迷航,並曾在多個歐洲國家報告過,包括荷蘭、英國、直布羅陀和捷克[55][56][57][58]。其常規繁殖範圍已不再向西延伸至德國,但仍偶爾有鳥隻出現在該地,每十年有幾次記錄。年輕鳥隻也廣泛擴散;德累斯頓自然歷史博物館有一個標本(C 21845),於1914年11月在薩克森州貝恩斯多夫附近被射殺。這是一隻幼鳥,雖然無法確定其確切年齡,但其身上有大量斑點,可能不超過20個月大[59]。
此外,在非洲也有迷航記錄,包括摩洛哥、阿爾及利亞、突尼西亞、利比亞、喀麥隆、乍得、肯尼亞、坦桑尼亞、贊比亞和博茨瓦納[21][60][61][62][63]。它有時也會出現在中亞和東亞的阿富汗中部和東部[64]。它們也可能出現在東亞,遍布俄羅斯遠東地區的南部、東部的中國,偶爾會在從緬甸和泰國延伸至馬來半島的東南亞地區[1][65]。偶爾在印尼(如蘇門答臘)也有記錄[66]。
越冬範圍
[编辑]專門的越冬地區通常比遷徙期間的範圍更有限和孤立[1]。主要的越冬地區集中在地中海盆地、中東和東洋界[6][67][68]。小區域可能存在於西南西班牙和毗鄰的葡萄牙、法國南部、意大利東北部、希臘西部(有時被認為是最常見的越冬雕種)、保加利亞南部、東部羅馬尼亞和摩爾多瓦南部的小區域[1][21][69][70][71][72]。其他越冬地區包括埃及東北部、蘇丹南部和毗鄰的南蘇丹、埃塞俄比亞中北部及中東的零散地區,包括北部以色列、科威特和中部敘利亞[1][21][73][74]。它們更連續地分布在阿拉伯半島南部沿海的大部分地區,包括沙烏地阿拉伯沿紅海海岸廣泛分布、也門西部和南部、阿曼南部、阿聯酋沿海及沙烏地阿拉伯東部。此外,它們在土耳其東南部、亞塞拜然、喬治亞東南部、伊拉克東部、伊朗西部、北部和東部廣泛分布,土庫曼斯坦南部、阿富汗西部和巴基斯坦最西部[1][75][76][77][78]。它們也零星地分布在巴基斯坦東部、印度北部、孟加拉、不丹南部及緬甸西北部。在印度,越冬範圍分布於印度河-恆河平原至比哈爾邦、賈坎德邦、西孟加拉邦至阿薩姆邦(包括北卡查爾丘陵),以及向南延伸至中部印度的東北丘陵州。它們曾經在馬拉巴爾海岸和卡納提克地區相對常見,但可能僅限於一百年前[6][15][79][80]。之後,它們在緬甸中部和南部、泰國中部和南部、寮國南部、越南北端、越南東南部零星地區和柬埔寨的大部分地區以及馬來西亞南部沿海分布[1][81][82][83][84]。在中國,越冬的花鵰從江蘇和安徽連續分布到廣東北部及台灣,偶爾出現在韓國[1][85][86]。
棲地
[编辑]
花鵰棲息在開闊的濕森林和森林邊緣,通常毗鄰沼澤、濕地、樹沼或濕草甸,以及河谷森林和泛濫平原森林[5]。與小斑鵰相比,它們通常更常見於濕地,但在中亞的乾燥丘陵森林中也可見到[5]。這兩個物種在棲地偏好上的差異在波蘭東北部得到證實,其中花鵰在泛濫頻率明顯高於小斑鵰的泛濫平原林地區築巢[87]。
雖然在受人類大規模開發影響的地區繁殖時通常較為稀少,但在愛沙尼亞也曾觀察到它們在耕地上狩獵,並在捷克低地農場上遷徙[5][58]。在俄羅斯,它們棲息於針葉林和開闊草原的過渡區(通常圍繞河谷),松樹林地、矮樹林附近、草原濕地和森林沼澤地區[48]。在哈薩克斯坦,它們的主要棲地是低地草原和森林草原交錯地帶的河岸森林[49]。
在冬季,花鵰的棲地與繁殖期間相似,通常出現在比其他鷹類更為潮濕的棲地,包括有林河口、紅樹林、沼澤、湖岸,尤其是在印度的湖區[5]。然而,花鵰也曾在東北非洲的半乾旱相思樹屬 稀樹草原中被記錄[5]。據報在厄立特里亞,它們出現在開闊的高原、村莊周圍和低地草原,而在蘇丹,它們通常出現在灌木叢地區[88][89]。在土耳其安卡拉冬季棲息的一隻花鵰,出現在高地森林地區[90]。在地中海盆地的一項研究發現,花鵰冬季最喜歡的棲地是鹽沼和含有淡水區域的沿海瀉湖[68]。在亞洲的冬季,它們並不罕見於稻田和有時的垃圾場,在這個季節中,它們對人類改變的地區更具適應性,儘管大體上仍偏好各種濕地、泥灘、大河、河口和紅樹林[30][15][84][91]。在阿拉伯地區,它們現在主要棲息於人造棲地,如污水處理場、水庫和農田,因為曾經圍繞沿海灣的原生紅樹林和蘆葦屬 蘆葦灘幾乎已完全消失[92]。在伊朗南部,它們通常出現在紅樹林區域[93]。在伊拉克,美索不達米亞沼澤是其重要的棲地[94]。在以色列,它們的冬季棲地是濕度最高的山谷和潮濕的開放區域,主要是靠近樹叢的耕地和魚塘,阿曼的棲地與此類似[75][92]。
花鵰通常分布於海平面到17,276英尺的高度,通常是低地鳥類。然而,在伊朗北部,它們已被記錄在高達17,277英尺的高度[5]。在拉達克的喜馬拉雅山上,也有一隻花鵰被記錄在17,278英尺的遷徙途中[30]。
遷徙
[编辑]

注意淺色的翅膀條紋。
花鵰幾乎完全是候鳥。然而,與其他猛禽相比,它們並不被認為是長距離遷徙的鳥類[95]。它們在八月底至十月間遷徙,有時會持續到十一月。回程通常在二月初開始,高峰期在三月,四月結束[5][21][58][96]。然而,在土耳其博斯普魯斯海峽附近,遷徙一直記錄到五月[97][98]。平均而言,遷徙高峰在其分佈範圍的東端更早出現,例如在不丹,最大的數量出現在二月下旬[99][100][101]。它們遷徙的時間比小斑鵰晚約兩週,回程時間也比小斑鵰早。在已知的遷徙中途停留點,小斑鵰的數量幾乎一般超過花鵰[8][30][102]。
在歐洲繁殖的花鵰可能遷徙至法國南部(尤其是卡馬格)、西班牙、意大利,有時到達瑞典。西部繁殖的鳥類也經常遷徙至北非,其中一些在摩洛哥、埃及、尼羅河谷、蘇丹、埃塞俄比亞,偶爾會更遠南[5]。來自不同起源地的鳥類可能會遷徙到中東(主要是阿拉伯地區)、南亞(從巴基斯坦,通常是旁遮普和信德省、印度北部和尼泊爾),向東到印度支那、馬來半島和中國南部及東部[5]。主要沿東歐軌跡遷徙的花鵰多數遷徙至中東或東北非洲,而其他則通過喀爾巴阡山脈遷徙至巴爾幹半島,有些繼續經由中歐和西歐遷徙至西南歐[95]。亞洲種群的主要越冬地位於阿拉伯半島、印度次大陸、中南半島和華東地區[95]。
在遷徙期間,花鵰通常每天遷徙約150 km(93 mi),但有時一天內可達到350 km(220 mi)。[103][104] 該物種的遷徙速度在貝加爾地區被記錄為26.6—45.5 km/h(16.5—28.3 mph),且高峰遷徙時間為中午至下午6:00。[105]
關於不同年齡和性別的遷徙差異的信息有限。相比於秋季遷徙,春季遷徙的數量通常較少。[8] 一般來說,該物種的遷徙以單隻或成對的形式在寬廣的前線移動,但曾在2月底於不丹北部觀察到多達10隻的群體。[5] 在馬來西亞,未成熟鵰的數量是成年鵰的六倍。[106] 在貝加爾湖,96%的遷徙花鵰被觀察到是成年鵰,這種不平衡引起了研究人員的擔憂。[105] 花鵰在傳統的遷徙瓶頸地區(如博斯普魯斯和紅海海峽)通常較少見。[5][97] 早期聲稱在博斯普魯斯秋季遷徙中有多達一千隻鵰的報導可能是錯誤的(雖然歷史最高遷徙數量可能已減少多達75%)。[8] 在俄羅斯伊爾庫茨克州的南貝加爾,秋季遷徙中花鵰僅佔觀察到的猛禽的0.2%(每年7–34隻;8年間共137隻)。[105] 最大的(現代?)計數是在埃及蘇伊士,秋季和春季分別記錄到86隻和74隻,另有少量記錄顯示在曼德海峽穿越進入非洲,儘管在北以色列秋季記錄的最高數量為85隻。[96]
一隻在10月底於沙特阿拉伯西部麥加附近捕獲的成年鵰被無線電追蹤到它飛行了850 km(530 mi)到達也門,並從11月底一直停留到2月初,隨後返回5,526 km(3,434 mi),經過伊拉克南部,越過伊朗高原,沿咸海南緣繞行,最終飛回其西伯利亞的繁殖區,靠近鄂木斯克,在不到一個月的時間裡完成了4,516 km(2,806 mi)的返程。[5] 對於一隻在愛沙尼亞繁殖的鵰進行的衛星追蹤確認了它在連續七年中穩定使用了西班牙加泰羅尼亞沿海的同一冬季棲地。[96] 儘管有些個體對冬季棲地表現出忠誠度,但一隻最初在阿聯酋捕獲的無線電追蹤個體在隨後的冬季被發現改道至巴基斯坦,而不是返回其哈薩克繁殖地,顯示了在這方面的某些變異性。[107]
一隻在沙特阿拉伯西南部過冬的花鵰(來自西西伯利亞的繁殖區)在冬季的平均家域為65 km2(25 sq mi),這一範圍在春季遷徙前縮小了24%,遷徙距離達到5,500 km(3,400 mi),[108] 而在西班牙的冬季領地則更小,僅有27.2 km2(10.5 sq mi)。[109]
花鵰的最南端遷徙記錄是一隻從波蘭別布扎國家公園遷徙了9,270 km(5,760 mi)到達非洲南部的贊比亞。[110] 其他幾隻據稱為花鵰的個體被追蹤到非洲的幾個地區,但其中近半數實際上是與小斑鵰的混種,並以該物種的更典型方式和地點進行遷徙。[104]
令人難以置信的是,至少有七個記錄顯示未成熟的花鵰在沙特阿拉伯過夏。[98] 類似地,有記錄顯示該物種在馬來西亞半島持續留存至至少5月。[84]
食性生物學
[编辑]
花鵰是一種略微機會主義的捕食者,但偏愛相當特定的獵物種類。[6] 它們的飲食主要由小型哺乳動物組成。[5] 儘管與小斑鵰有一些飲食相似性,但花鵰的飲食中包括更多鳥類,且偏好較大的獵物。[5] 除了哺乳動物和鳥類外,花鵰有時會捕食兩棲動物、爬行動物(主要是中小型蛇類),偶爾還會捕食小魚和昆蟲。[5][25] 食丸被認為是確定花鵰飲食最可靠的方法,因為僅靠獵物殘骸分辨可能會偏向鳥類。[30]
如其棲地偏好所預期,花鵰傾向於捕食與水相關的脊椎動物。[5][21] 一般來說,其獵物組成相對多樣,已知的獵物種類略少於150種;這比亞洲草原鵰的已知飲食更為多樣,與小斑鵰的飲食相似,且可能約為白肩鵰飲食多樣性的二分之一。[7][6][111][112]
花鵰主要在空中狩獵,在相對開闊的地面上飛行(有點像鷂)或在高空滑翔,當發現獵物時急速下落或俯衝。[5][6] Brown和Amadon描述了正在狩獵的花鵰,說道:「儘管它不是一種非常活躍的物種,但也不能說它是遲緩的,在空中它看起來像一隻真正的鵰。」[8] 花鵰會通過低空飛越水鳥群來驅散它們,然後選擇孤立的個體進行攻擊。[5] 有時,花鵰會從棲息處靜候獵物,這種方法更常見於其他分布範圍相似的鵰,並且經常在地面上狩獵。它大多在地面或水上捕食。[5][6] 該物種有時也會對其他猛禽進行盜食寄生攻擊。[5] 儘管花鵰幾乎全年都會食腐,在非繁殖期,這種情況可能更為普遍,因為此時老鷹大多將新鮮的獵物帶回巢中。[7][6] 與亞洲草原鵰等其他捕食者一樣,花鵰在非繁殖期經常被草原野火和成群的蝗蟲所吸引。。[5]
在波蘭別布扎國家公園的繁殖季節中,研究了花鵰的狩獵行為。研究發現,飛行和狩獵的高峰時間為上午10:00至下午2:00,鵰很少在上午9:00之前飛行,並且經常在下午2:00到4:00之間休息。雄鵰(主要的食物供應者)可能提供的獵物比季節後期雌鵰恢復狩獵時更加多樣化。在別布扎,狩獵領域受到來自同種、小斑鵰及其他大型猛禽的保護。根據別布扎的數據,雄鵰的狩獵成功率在7月中旬前為34%,此後下降至20%。[113] 花鵰的狩獵成功率似乎相當高——上述在繁殖季節大部分時間內的34%遠高於金鵰(約20%)、小斑鵰(24%)和白腹隼雕(28.5%)的狩獵成功率。[114]
有時據說它們捕食的獵物大多僅重達250 g(8.8 oz)。[5] 其中一個來源估計,它們飲食中的大約22%獵物重量為63 g(2.2 oz)或以下,37%的獵物重達63—125 g(2.2—4.4 oz),約30%重達125—250 g(4.4—8.8 oz),總體來說大多數獵物重於1,000 g(2.2磅)。根據此來源,花鵰的平均估計獵物大小可能約為157 g(5.5 oz)。[10]
白俄羅斯的一項大規模研究發現,花鵰的飲食主要由體重為51—200 g(1.8—7.1 oz)(41.9%)和11—50 g(0.39—1.76 oz)(38.3%)的獵物組成。[111] 然而,對於總體獵物生物量的最大貢獻來自體重為601—1,200 g(21.2—42.3 oz)(34%)的獵物。[111] 在這項研究中,雄鵰交付的獵物平均估計大小為161.2 g(5.69 oz)。[111] 相比之下,一項愛沙尼亞研究估計的數值僅為57.8 g(2.04 oz)。[115] 平均獵物大小大致與較大的草原鵰相似,且稍高於小斑鵰,後者傾向於集中捕捉體重低於63 g(2.2 oz)的獵物(約占飲食的60%);然而,大多數鵰的典型獵物(白肩鵰)的重量比花鵰的典型獵物稍高至顯著高於(如金鵰)。[10] 儘管如此,有時花鵰被認為能成功捕獵大型獵物。[6][21]
在已知最大規模的花鵰食物研究中,該物種的飲食在白俄羅斯波利西亞的三種不同棲地中進行了研究,從自然到混合再到改良棲地。共識別出797件獵物(但通常未標識到種級),其中包括哺乳動物(40.4%)、鳥類(36.0%)、無脊椎動物(15.8%,主要是甲蟲)、爬行動物(5.7%)、兩棲動物(1.3%)和魚類(0.9%)。各類田鼠是重要的食物來源——即普通田鼠、苔原田鼠和東歐田鼠——合計占飲食的23.4%。其他重要的獵物包括水䶄(8.9%)、丘鷸(4.1%)、綠頭鴨(3.3%)、水雞(3.1%)、未確定的小型雀形目鳥類(2.9%)、斑胸秧雞(2.6%)和水游蛇(2.3%)。在生物量中顯著但數量較少的有北白胸刺蝟、歐洲鼴鼠、涉禽鴨類、蒼鷺、大麻鷺和黑琴雞,少量捕獲了非常大型的鳥類。[111]
另一項研究中,在白俄羅斯斯摩棱斯克湖國家公園記錄的102件獵物中,最常見的是歐洲水田鼠(38.2%)、歐洲鼴鼠(7.8%);5.9%各自為鼩鼱、田鼠和未確定的雀形目鳥類;以及蛙(8.7%)。[116] 在愛沙尼亞的一個巢中,105件視覺識別出的獵物中,田鼠屬(Microtus) 物種和進一步未確定的囓齒動物約占數量的63%但僅占生物量的28%,而鳥類占數量的19%但占生物量的56%;45%的鳥類獵物物種是中型鳥類,如花尾榛雞、灰山鶉、小辮鴴和黑頭鴉。[115]
其他歐洲研究主要集中在冬季的花鵰。 在希臘阿姆弗拉基亞濕地,共確定95件獵物,幾乎完全由水鳥組成。這裡的主要獵物有小水鴨(數量占17.9%,生物量占15.9%)、紅冠水雞(16.8%,14.9%)、赤頸鴨(11.6%,27.4%)、未確定的鴨屬 (Anas) 鴨類(5.3%,7.8%)、白冠雞(4.2%,8.7%)和小白鷺(3.2%,4.7%),還有少量的步行蟲科甲蟲、雀形目鳥類和蛇類。[117]
在西班牙阿利坎特省的埃爾豐多自然公園進行的為期8年的研究中,幾乎全部捕捉的是大型獵物,幾乎沒有小型囓齒動物(如田鼠)。在發現的100件獵物中,主要的獵物是紅冠水雞(數量占23.1%,生物量占15.2%)、小水鴨(8.97%,6.44%)、黑鼠(7.69%,3.01%),以及未確定的大鼠屬(7.69%,2.76%)。其他經常捕捉的獵物包括紅嘴鷗、歐亞白冠雞和小辮鴴。在生物量中占大部分的大型獵物是鯉魚(占生物量的18.9%)、蒼鷺(11.7%)和穴兔(9%)。[118]
在俄羅斯西部的巢中,發現飲食中平均有53%是小型哺乳動物,45%是鳥類。[115] 在列寧格勒地區的一項研究中,花鵰的飲食由79件獵物組成,主要是水䶄(占51%),其次是引進的麝鼠(3.9%)和小水鴨(3.8%),還有蛙類(20.2%),很可能是林蛙和一些田野林蛙。[119]
在別拉亞河地區發現322件非腐肉的獵物,59%為哺乳動物。飲食主要由歐洲水田鼠組成,占32.6%,其次是小型田鼠和老鼠。研究發現,別拉亞地區的花鵰攝食了大量的爬行類獵物(占19.5%)——事實上,有15%的脊椎動物獵物是極北蝰。花鵰捕捉了中等大小的蛇,但並未見到捕食小蛇,也沒有捕食許多特別大的蛇;牠們經常被發現抓住蛇的頭部。在別拉亞地區,鳥類僅占飲食的6.5%,主要在奧卡自然保護區有顯著地位。該地區的花鵰有時也參與食腐,包括駝鹿屍體。[120]
在來自伏爾加地區、烏拉爾山脈和西西伯利亞的綜合研究中,花鵰的飲食中有74.7%是哺乳動物,來自482件獵物。其主要獵物種類是歐洲水田鼠,平均占飲食的32.4%(28.1%-36.8%),其次是普通田鼠,平均占11.4%(0%-17%),苔原田鼠占6.2%(1.9%-16.9%),鳥類占16%,其中最重要的是白冠雞和鸊鷉屬鷿鷈,其次是秧雞。[7] 在圖瓦共和國,由於更偏向陸地的高地狩獵,據報導達烏爾鼠兔是花鵰的主要獵物。同樣,在哈卡斯,據說花鵰主要捕食長尾地松鼠,這是唯一已知在花鵰繁殖區中地松鼠比田鼠更受青睞的地方。[121]

在非歐洲的越冬區,花鵰的飲食普遍所知不多。與許多其他類似地區的猛禽一樣,遷徙的花鵰通常會禁食直到抵達其越冬終點站。[8][6] 在亞洲最受研究的與飲食行為相關的越冬族群可能位於印度婆羅多布爾縣的凱奧拉德奧國家公園。在這裡,花鵰佔據的棲地範圍比其他花鵰和鵰屬鵰類更大。花鵰在此不具領域性並自由活動,據記錄其利用當地9 km(5.6 mi)長的沼澤地。在這個地區,冬季花鵰的數量可達30隻,其數量在11月和12月達到高峰,然後在1月後減少。與此地其他越冬的遷徙猛禽一樣,例如鸢,花鵰成為高度機會主義的捕食者,偏愛容易獲得的食物。牠們經常食腐,搶奪其他鳥類的食物,捕食擱淺的魚,並且可能最經常捕食年幼的鷺、鸛和其他水鳥,這些鳥類來自鷺鷥林。在捕食這些鳥類時,花鵰通常會懸停以打亂鷺巢群,然後當鷺巢群穩定下來時,突然俯衝到目標巢穴上,拔掉巢中的雛鳥(年幼的水鳥)羽毛,並開始進食。在婆羅多布爾的花鵰對於動作緩慢的獵物略有偏好,但也會捕捉飛行迅速的鳥類如涉禽和鴨。鵰會飛低掠過水面不斷“嗡嗡作響”以測試白冠雞群,直到能攻擊一隻孤立的個體。牠們經常棲息在最喜愛的瞭望點上,在婆羅多布爾最喜歡阿拉伯膠樹。儘管牠們通常在白天狩獵,但在婆羅多布爾已推測可能存在夜間狩獵。在婆羅多布爾的惡劣天氣期間,花鵰可能會暫停狩獵。多達7到10隻花鵰會因同類的鷺巢群獵殺而被吸引,隨後經常互相推擠。這常常導致鵰不小心將獵物掉入水中,儘管掉在乾地上的獵物經常成為野豬和豺狗的食物。曾有一例,兩隻成年花鵰和一隻幼年白肩鵰在一天內毀壞了至少30個水鳥巢穴。在對花鵰的79小時觀察中,牠們49%的時間用於覓食,隨著3月份食物變少,這一比例上升至72%。同時,牠們26.4%的時間用於休息,20.6%的時間用於翱翔,而在2月份翱翔時間增加到35%。花鵰每日平均消耗240 g(8.5 oz)食物,大多數食物可以養活牠們幾天。據報導,牠們捕獵的獵物種類是該地區猛禽中最多樣化的。[6][122][123][124][125]
在整個印度次大陸,大花鵰已知會自由地拾食腐肉,並且捕食青蛙(尤其是虎紋蛙)、變色龍、樹蜥屬蜥蜴、蛇、囓齒動物和小型哺乳動物。據報導,該地區大花鵰對鳥類獵物有一般的捕食傾向——主要是較大的秧雞類,例如黑水雞、白冠雞和灰頭紫水雞,以及水禽和(多為幼年的)鸛、鷺和白鷺;然而,丘陵地區的鳥類,如棕腹樹鵲、灰斑鳩和西棕胸佛法僧似乎也被包括在內。儘管很少在大型腐肉處出現,但印度次大陸的大花鵰似乎會被玉帶海鵰、白兀鷲和黑兀鷲弄傷或部分吃掉的泥龜吸引。[6][30][126]
.jpg)
在泰國中部平原,獲得了一些關於冬季大花鵰飲食的軼事證據。在這裡,牠們被觀察到吃乾涸池塘中的死魚,也會主動捕食和掠奪其他猛禽的食物。牠們被看到捕食與大型農場鴨群分開的家鴨,以及吃被毒藥殺死的樹鴨屍體,這些毒藥本來是用來殺死蝸牛的。[82] 根據報導,稻田鼠似乎是該地區大花鵰的重要獵物之一。[5][30] 儘管通常不被認為具備攻擊大型或多樣化獵物的能力,有時大花鵰似乎能捕捉到非常多樣且有時體型相當大的獵物。[6][10][113]
已觀察到有時會捕捉小型無脊椎動物,包括步行蟲、蝗蟲和非本地的克氏原螯蝦。[111][118] 大花鵰捕捉的鳥類範圍廣泛,從重量僅18.4 g(0.65 oz)的蘆鵐到成體大白鷺、蒼鷺、野鴨和幼年的鸛類如白頭䴉鸛和鉗嘴鸛等體型較大的水鳥。[6][29][111][118][127] 有時大花鵰會攻擊或拾食鶴類,但據報導許多攻擊嘗試並不成功。曾有報導觀察到花鵰拾食灰鶴(Grus grus)屍體,以及對成體蓑羽鶴(Grus virgo)的捕食失敗案例。花鵰也被認為對丹頂鶴(Grus japonensis)的幼鳥構成潛在威脅,而丹頂鶴則以激烈的方式保護其雛鳥。[111][128][129]
除了鳥類獵物外,哺乳動物的捕食範圍從巢鼠和普通鼩鼱,重量不超過8 g(0.28 oz),到可能重達3,000 g(6.6磅)的歐洲野兔。有時大花鵰可能會捕食約半打的鼬科物種,主要是各種黃鼠狼和白鼬,但也包括較大的物種如水鼬和貂。[113][111][130][131]
種間捕食關係
[编辑]
花鵰經常與其繁殖區和越冬區的多種相似鷹類物種有廣泛的重疊。[5][25] 在其繁殖範圍的較大部分,花鵰與小斑鵰(Clanga pomarina)是異域物種形成的。[42] 當牠們重疊時,牠們有著相似的飲食,但花鵰傾向於專注於水生物種,並捕食相對較多的鳥類,而小斑鵰則通常專注於田鼠、小蛇(如水游蛇)和青蛙。[132] 因此,小斑鵰傾向於在稍微乾燥的環境中築巢,通常稍微遠離濕地和氾濫平原,更容易適應人類開發的碎片化地區。[9][87][133] 在中心分佈區域內,較為相似的是白肩鵰(Aquila heliaca)和亞洲草原鵰(Aquila nipalensis)等較大型鷹類。[5][8]
此外,這些物種進行的大致相似的遷徙路線,儘管亞洲草原鵰是最常見且規律的遷徙者(儘管其數量大幅減少),從非洲到南亞都可以見到牠們,而花鵰和白肩鵰則會定期出現在中東,並且在非洲的數量很少見。[134][135][136] 然而,花鵰顯然與其他鵰類有所區別,因為牠偏好濕地和部分林地的棲地以及其中的獵物。白肩鵰也在林地築巢,但通常在山區,牠偏好社交或獨居的陸生哺乳動物和鳥類,包括野兔、倉鼠、地松鼠和刺蝟,以及雉、鴉科和其他中型鳥類。[137][138] 與此同時,亞洲草原鵰偏好通常較乾燥且非常開放的草原棲地,通常在平坦、稀疏的棲地中選擇一個高地或岩石築巢,並且非常喜歡地松鼠,並以其他小型陸生物種如達烏爾鼠兔、田鼠和鼢鼠作為補充。[139][140] 這些棲地通常在築巢時將這些鵰類與大花鵰分開;然而,在印度、地中海盆地和中東等越冬區域,卻會出現顯著的交集。[6][122][127][141]
所有這三種鵰在冬季都是公認的不挑食機會主義者和拾荒者。牠們會自由地接近人類垃圾(偏好家畜屍體堆放處)、撿拾未被佔有的腐肉、搶奪其他猛禽的獵物、殺害水鳥等獵物的幼鳥、尋找昆蟲群或大量出現的昆蟲(亞洲草原鵰比其他鵰更如此),並跟隨草地火災。在這三種鵰中,亞洲草原鵰在冬季通常是最不積極捕獵的,白肩鵰則是最有可能繼續狩獵活物(並且最常棲息),而花鵰的行為則介於兩者之間。花鵰是三者中最不可能去尋找腐肉或屍體堆放處的,但在印度次大陸,牠們都會大量分享食物來源,如水鳥的雛鳥。[5][6][122][123] 當衝突發生時,鵰的體型決定了其在階級中的地位——白肩鵰是最具優勢的,其次是亞洲草原鵰,而花鵰在兩者面前則顯得較為服從。在婆羅多布爾進行的研究中,這三種鵰與短程遷徙的玉帶海鵰競爭,玉帶海鵰與白肩鵰爭奪主導猛禽的地位,而所有較大的鵰類則壓倒了較小的、定居的印度花雕。[123] 與冬季花鵰經常相關聯的非鵰猛禽黑鳶也被相似的食物機會吸引。[142]

在拾荒時,如果舊大陸鷲類存在,花鵰通常不會靠近腐肉。[6] 花鵰在冬季期間是出色的劫掠者。牠們經常搶奪各種其他猛禽的食物,包括黑鳶、鶚、西方和東方澤鵟,甚至其他鵰類,包括像白腹海鵰和亞洲草原鵰這樣的較大物種。儘管花鵰通常在牠們面前處於劣勢,但在婆羅多布爾,花鵰常常成功地將亞洲草原鵰從有爭議的食物中趕走,而亞洲草原鵰在飽餐後顯得更加柔順。牠們通常在其他猛禽起飛時搶奪其獵物,利用對方試圖保持平衡的時機,快速奪走獵物並迅速逃走。[6][82][122][126][143]
更不常見的是,花鵰有時會將其他猛禽作為獵物,這些包括黑鳶、靴鵰、西方澤鵟和歐亞鵟,以及一些貓頭鷹,如長耳鴞和短耳鴞。此外,牠們還被認為是兀鷲幼雛的潛在捕食者。[6][111][118][144]
花鵰本身很少有被充分記錄的捕食者。[8] 雖然這可能部分是由於研究不足,但作為一種相當大且強大的猛禽,牠們通常扮演頂級捕食者的角色。[25][130] 儘管如此,一個被充分記錄的花鵰掠食者是雕鴞。[145][146] 此外,歐洲松貂也被發現會捕食花鵰的幼鳥。[113]
繁殖
[编辑]花鵰通常成對或單獨出現,但在冬季,有時會在有吸引力的食物來源附近形成小至大型的群體。[5] 這種鵰通常在遷徙過程中單獨出現,儘管有時也會成對、三隻或更多一起出現。[5]
在大部分地區,這種鵰從4月底至8月間進行繁殖。[5] 然而,在巴基斯坦和印度次大陸,牠們的繁殖期根據不同的報告顯示為11月至3月,有時甚至延伸至6月至7月,顯示出那裡的繁殖時間表並不一致。[5][6]
這種鵰的求偶行為尚不為人所知,但包括單獨或互相在高空盤旋,雄鵰向雌鵰俯衝,雙翼半閉,並伴隨著大量鳴聲。[5] 領域範圍在理想區域內可達15至30 km2(5.8至11.6 sq mi),通常在保護區範圍內,然而在其他地方則要大得多。[30]
過去有報導稱,大花鵰的巢可彼此相距僅100米(330英尺);在某個案例中,四對鵰在一個僅有0.6 km2(0.23 sq mi)的區域內築巢。[8] 更典型的範圍可能是在40—52 km2(15—20 sq mi)的區域內。[8] 在伏爾加河–烏拉山脈地區的森林區域中,大花鵰的密度為每100 km2(39 sq mi)有4.76對(根據確認數量)至6.15對(根據可用棲地的預測)。而在西西伯利亞地區,則為每100 km2有6.55至8.76對。在伏爾加河–烏拉山脈區域,當地優選的棲地(如被淹的赤楊)可容納高達3.58至17.01對。該地區巢穴之間的平均距離為7.3 km2(2.8 sq mi)。[7] 在哈薩克斯坦的伊希姆河流域,每100 km2有0.54對。100米(330英尺) 而在哈薩克斯坦的額爾齊斯河松林中,內部森林邊緣的密度為每100 km21.08對,森林邊緣沿湖泊和沼澤地區的密度則高達每100 km213.23對。
哈薩克斯坦密度最高的區域可能是伊希姆河流域,該地區可能擁有全國39%的繁殖對。[147] 此外,在適合的俄羅斯棲地,據說(至少在歷史上)每1.6 km(0.99 mi)的河岸都可以找到巢穴,而且繁殖對在隨後幾年內通常會重複使用這些巢穴。[8]
巢
[编辑]這種鵰會建造大型樹枝巢,巢穴直徑可達70—110 cm(28—43英寸),深度可達100 cm(39英寸)。[5] 在印度次大陸,巢穴顯著較小,直徑約為60 cm(24英寸),短於鵰本身的全長,深度僅為5—15 cm(2.0—5.9英寸)。[6]
不同尋常的是,這些鵰通常使用仍帶有葉片或綠色針葉的新鮮樹枝來建巢(大多數其他隼形目鳥類偏好使用稀疏或無葉的樹枝建巢)。[7] 巢穴內鋪有綠葉、針葉和草,這是隼形目鳥類的常見做法,並且可能會在整個繁殖期不斷添加。[5][8] 巢穴通常位於樹上,通常在主叉、大側枝或大闊葉樹的頂部,靠近森林邊緣。[5][7]
俄羅斯的綜合研究發現,約68.7%的大花鵰巢穴位於闊葉樹上,其餘則位於針葉樹上。[7] 在俄羅斯的下諾夫哥羅德地區,11個觀察到的巢穴中有10個位於樺樹上,一個位於普通赤楊上。[148] 在波蘭,樺樹(如毛樺)似乎很受歡迎。[113] 在伏爾加河–烏拉山脈地區,赤楊林是首選,有71.4%的繁殖對使用了已被發現的活躍巢穴,而在西西伯利亞地區,則偏好松樹林(55.9%)。[7] 在阿爾泰-薩彥山脈地區,首選的築巢樹木是樺樹(50%)和落葉松(31.25%)。[121] 據報導,在巴基斯坦,曾使用阿拉伯膠樹和芒果樹築巢;在印度則用Mitragyna parvifolia ,據報導,有時甚至在農田上築巢。[6][30]
俄羅斯的研究發現,巢幾乎一般在氾濫平原的森林中。[7][121][148] 巢可以位於距地面或水面5—25米(16—82英尺)高的地方,雖然通常是8—12米(26—39英尺)。[5] 在伏爾加河–烏拉爾山脈地區,巢位置距離最近的森林邊緣平均為510.7米(1,676英尺),但往往位於森林最密集的部分。[149] 從伏爾加河–烏拉爾山脈地區的83個巢樣本中,巢的平均高度為9.58米(31.4英尺)。[7] 在下諾夫哥羅德,巢穴的高度為距地面7—14米(23—46英尺)。[148] 在西西伯利亞,巢穴高度範圍從3—20米(9.8—65.6英尺)不等,平均為7.22米(23.7英尺),並且在以針葉林為主的西西伯利亞地區,巢穴位置顯得較低。[7] 在阿爾泰-薩彥地區,巢穴高度更低,平均為6.5米(21英尺)。[121]
巢很少出現在無樹地區的灌木中。[5] 據報導,阿爾泰-薩彥地區的一個巢穴僅位於距地面2米(6.6英尺)的地方。[121] 更罕見的是,據報導,有些巢穴甚至位於地面上。[30] 有時,它們可能會使用其他鳥類的巢穴——最有可能是其他猛禽的巢穴,但據報導,有一次甚至使用了一個歐亞喜鵲的巢穴。[30][150]
幼鳥的發育
[编辑]
在俄羅斯,花鵰據報導通常不會在5月前產卵,但有時會早至4月底開始產卵,西部地區的產卵時間類似(但略早一些)。[8][30] 一窩通常有兩顆蛋,但有時巢中會有一到三顆蛋。[5][8] 蛋呈灰白色卵形,通常無光澤且無斑點。然而,有時它們可能會出現一些深棕色斑點或斑塊以及稀疏的灰色殼痕。[8] 蛋的高度可能範圍在63至74 mm(2.5至2.9英寸),直徑範圍在47.8至56.7 mm(1.88至2.23英寸)之間。[7][8][151]
大多數資料指出孵化期為42–44天,但在南部白俄羅斯,曾記錄到孵化期僅為39天。[5][152] 孵化從第一顆蛋開始。[8] 曾經認為雄鵰不參與孵化(通常,正如許多猛禽一樣,牠們主要負責獵物的運送)。[8] 然而,在上述的白俄羅斯研究中,雄鵰在白天平均孵化57.3分鐘。[152]
在阿爾泰山脈的針葉林地區,六個花鵰領域中,每個成功的巢穴平均有1.33隻幼鳥,或每個占領的巢穴有1.0隻幼鳥。兩隻阿爾泰花鵰領域位於白肩鵰的廢棄領域上。[153] 與此同時,在伏爾加河–烏拉爾山脈和西西伯利亞地區,平均幼鳥數分別為1.24和1.42。[7]
當第二隻幼鵰孵化時,大花鵰的幼鳥之間的體型差異明顯,較年幼的通常會死亡,往往是通過同胞殺害。[8] 猛禽,尤其是鷹類之間的競爭經常導致年幼幼鳥因飢餓或被較大幼鳥故意殺害,這並不罕見,通常被假設為一種保險機制,較年幼的幼鳥在較大幼鳥意外死亡時起保險作用;否則,較年幼的幼鳥(不一定是健康狀況不佳)可能會預期死亡。[154][155] 然而,這種鵰比小斑鵰更常見的是能夠撫養兩隻幼鳥至離巢。[8][156] 然而,在奧卡自然保護區的50個巢穴中,僅有一對在一年內成功撫養了兩隻幼鳥。[157]
在波蘭的一項實驗中,為了拯救較年幼的幼鳥,將其從巢中取出。當時,較年幼的幼鳥重310 g(11 oz),而較大幼鳥重1,050 g(2.31磅)。在巢外,該幼鳥在人類最小干預的情況下被養育,以避免銘印,除了人工餵食之外。該幼鳥與一隻白肩鵰和一隻小斑鵰共享籠子,這兩隻鵰對它都漠不關心,既不照顧也不餵食它。在幼鳥準備離巢時,該幼鳥被成功重新引入到親鳥的巢中,離巢並最終獲得獨立。[157]
母花鵰在孵蛋時會緊緊守護巢穴,但在受到干擾時可能會離巢一整天才返回。[8][152] 到7月底至8月初,幼鳥已完全長出羽毛,並在五天內首次飛行。[8] 幼鳥在60至67天後離巢,平均約為62天。[5] 在阿爾泰-薩彥地區,每個成功巢穴的平均離巢幼鳥數在圖瓦為1隻,而在哈卡斯為1.8隻。[121] 在哈薩克斯坦的研究中,11個巢穴平均產生1.38隻幼鳥。[147] 在西西伯利亞,66個巢穴平均產生1.44隻幼鳥。[158]
幼鳥在離巢後的30天內仍依賴父母。[5] 在分散前,年幼的花鵰可能會在當地的開闊草原中游蕩。[8] 一項在波蘭進行的研究發現,雌性花鵰通常在幼鳥之前2-3天離開巢穴。而雄性往往最後離開,大約在雌性之後一週。在波蘭的研究中,成鳥直接飛往博斯普魯斯,而幼鳥有時不那麼直接。[159] 大多數在9月底前離開波蘭。秋季期間,在波蘭的其他地方曾觀察到幼鳥在最終遷徙前游蕩。[159]
混種化
[编辑]曾經,花鵰和小斑鵰可能因棲地的不同(儘管可能接近)而大部分時間彼此隔離。[4] 在最後的冰河時期結束時(在某個全新世早期),氣候變化促進了森林的生長,這些森林取代了原來的草地邊界,使得兩種花鵰得以進入彼此的範圍。[4][160] 現在已知混種化現象廣泛存在。兩種花鵰在其重疊的範圍內均會出現混種,這個範圍大約有600,000 km2(230,000 sq mi)。混種現象在歐洲俄羅斯大多是推測得知,這裡是小斑鵰的東部極限,因此可能是最近才發生混種化的地區。[11][148][161]
這兩種花鵰的混種通常會顯示出頸背斑塊(純大花鵰沒有),翅膀上有中等數量的斑點,且通常體型比純小斑鵰大。[162] 儘管它們具有中間特徵且比小斑鵰體型更大,但花鵰的混種化被認為是花鵰領域被放棄的跡象,而這些領域被適應能力更強、數量更多的小斑鵰所取代,這一點在愛沙尼亞的一項研究中得到了證實。[11] 該研究發現,在愛沙尼亞,混種花鵰的數量是純花鵰的兩倍。[11] 在立陶宛,情況更為嚴峻,到了2000年代中期,已經找不到任何純花鵰配對,161隻繁殖的花鵰中只有2.7%是花鵰,其餘都是小斑鵰。[163] 估計立陶宛有大約1,000對繁殖中的小斑鵰,其中約有37對中有一隻配偶是花鵰。[163]
波蘭和愛沙尼亞的研究都顯示混種配對中的伴侶更替率可能很高,波蘭的數據顯示,大約71%的雄鵰在後續年份被補充。[11][164] 此外,波蘭的數據還顯示,混種花鵰偏好小斑鵰的棲地,這些地區遠離花鵰的濕潤棲地,且往往靠近人類的開發區,當地純花鵰配對數量減少了50%,而混種配對數量增加了30%。人為環境變更一般對小斑鵰有部分益處,但通常對花鵰有害。[164][165]
狀況和保護
[编辑]
儘管擁有相當廣泛的繁殖範圍——從波羅的海橫跨歐洲向東延伸至太平洋,並在印度次大陸有小範圍分佈——這種鵰的密度極低。[5] 在它分佈的幾乎每個國家中,[21] 花鵰都被IUCN列為易危的滅絕物種。[1]
過去對該物種的種群和趨勢研究較為匱乏,但已發現明顯的下降趨勢。[5][30] 1990年代的粗略估計顯示,在東北波蘭的一個約17.611平方公里的巨大區域內只有11對,在俄羅斯歐洲部分的一個17.612平方公里的區域內只有20至30對,烏拉爾以西不超過900對。[5] 隨後的更精細研究顯示,歐盟內的繁殖對數量為810至1,100對。[21]
此外,1990年代的印度越冬種群推測表明,東部種群的數量肯定不到四位數。[5] 國際鳥盟在1990年代估計俄羅斯的種群數量為2,800至3,000對。最近,國際鳥盟估計全球總成熟個體數量不超過3,800至13,200隻。[1][5][30]
顏色標記恢復研究
[编辑]一項顏色標記恢復研究發現,在1,370例歐洲花鵰的標記恢復中,只有3.6%是花鵰,混種花鵰和小斑鵰佔2.7%,其餘都是小斑鵰。[166] 花鵰作為繁殖物種已在匈牙利、羅馬尼亞、保加利亞、捷克共和國(在那裡它們可能從未穩定繁殖)、斯洛伐克以及以色列滅絕,最後一次繁殖是在1960年代。同時,芬蘭的繁殖種群也可能幾乎消失。[5][30][21]
估計烏克蘭的花鵰數量在1985年穩定減少至40–50對,且從1920年代到1990年代,烏克蘭的總數量減少了12–20%。[5][167] 在愛沙尼亞,僅從2004年到2010年,花鵰的數量就減少了14%,而且這種減少趨勢早已被觀察到。[12][168] 白俄羅斯擁有多達150–200對繁殖對(確認的數量略超過100對),被認為是俄羅斯以外最重要的繁殖區。[21]
歐洲俄羅斯的總數量在1960年代曾估計約為1,000對繁殖對,現在估計少於700對。[30][169] 該物種的分佈範圍在俄羅斯遠東地區縮小,曾經廣泛分佈的區域現在僅限於中阿穆爾河以下、沿著烏蘇里江以及南濱海邊疆區,儘管有轶事顯示,在從烏拉山脈到中鄂畢河的整個西西伯利亞低地區域,它們仍然相當常見。[30] 在哈薩克斯坦,估計有74–97對繁殖對。[147]
冬季估計數量較為分散,對印度的調查顯示,儘管它們繼續廣泛存在,但數量可能略有下降且分佈更為分散。[6][170][171][172] 在亞美尼亞,它被認為是該國30種猛禽中最稀有的兩種之一,與白肩鵰齊名。[173] 在地中海盆地的冬季數量約為300–400隻,其中略低於34%在以色列,略低於32%在希臘,16%在土耳其,羅馬尼亞和西班牙數量略少,在東南歐、蒙特內哥羅和法國的數量極少。[68] 其他來源顯示,土耳其的冬季數量約為50隻。[90] 該物種在衣索比亞和厄立特里亞出現相當罕見,大多數情況下都是單獨且零星出現。[73]
威脅與保護努力
[编辑].jpg)
該物種的主要威脅是棲地退化和棲地喪失。[1][30] 花鵰似乎對棲地變化高度敏感,尤其是濕地排水、農業活動強化和洪泛平原管理措施的放棄。[87][174]
不利的濕地管理過程還影響了該物種的冬季棲地,在沙烏地阿拉伯,至少部分影響已因該物種適應人造水域而被抵消(不過在繁殖期,尚無證據表明它們能很好地適應人造區域)。[175][176] 在泰國,可用的人造棲地因旱季稻田耕作的變化和城市化的逐步擴展而縮小,此外還可能受到滅鼠藥的使用和其他毒物的危害,這可能損害了能夠在那裡過冬的該物種數量。[82]
其他已知的威脅包括繁殖季節的人為干擾,如森林作業現已被確認為巢址主要的干擾來源。[30][174] 此外,花鵰還受到無意中的中毒和與人造物體(尤其是輸電線)碰撞的威脅。[5][177] 中毒已被確認是中國山西一個保護區的主要死亡原因,在那裡,花鵰會捕獵中毒或垂死的雉雞,隨後自己也會死亡,這成為當地死亡的主要原因。[30][178]
在馬來半島,繼1960年代至1980年代由於環境變化有利於鳥類食腐動物而數量短暫增加之後,隨後數量的劇減至幾乎沒有,可能是由於農藥和其他毒物的使用所致。[21][84] 花鵰的實際趨勢有時會被誤認所掩蓋。[5]
此外,如前所述,該物種面臨混種化的威脅,並最終可能被小斑鵰取代,因為其分佈範圍逐漸向東擴展。[4][11] 花鵰在只有少數國家中受法律保護,使得保護工作變得困難。[1][30] 受法律保護的國家包括白俄羅斯、愛沙尼亞、法國、希臘、拉脫維亞、波蘭、羅馬尼亞、俄羅斯和泰國(名義上)。21世紀已成立專門應對花鵰的工作小組。[1][30][179] 這些工作小組已在白俄羅斯、愛沙尼亞和烏克蘭——該物種在歐洲剩餘的核心繁殖區——開展保護工作,並成功制定了繁殖季節期間在巢址附近限制林業活動的措施。[1][30][179]
在下諾夫哥羅德,建造人工巢台似乎對花鵰幫助不大——不同於其他猛禽如魚鷹、白尾海鵰和金鵰——因為只有一對花鵰被記錄到使用人工巢台作為巢。[148][180] 值得正面肯定的是,通過微衛星研究發現,歐洲花鵰種群保留了相當高的基因多樣性,這意味著該物種面臨種群瓶頸的迫在眉睫的威脅不大。[4][181][182]
保护
[编辑]- 中国国家重点保护动物等级:一级
圖集
[编辑]
参考文献
[编辑]- ^ 1.00 1.01 1.02 1.03 1.04 1.05 1.06 1.07 1.08 1.09 1.10 1.11 1.12 1.13 1.14 1.15 1.16 1.17 1.18 1.19 1.20 1.21 1.22 BirdLife International. Clanga clanga. The IUCN Red List of Threatened Species. 2021, 2021: e.T22696027A203868747 [11 November 2021]. doi:10.2305/IUCN.UK.2021-3.RLTS.T22696027A203868747.en
 .
.
- ^ 2.0 2.1 中国科学院动物研究所. 乌鵰. 《中国动物物种编目数据库》. 中国科学院微生物研究所. [2009-04-04]. (原始内容存档于2016-03-05).
- ^ 3.0 3.1 3.2 Helbig, A. J., Kocum, A., Seibold, I., & Braun, M. J. (2005). A multi-gene phylogeny of aquiline eagles (Aves: Accipitriformes) reveals extensive paraphyly at the genus level. Molecular phylogenetics and evolution, 35(1), 147-164.
- ^ 4.0 4.1 4.2 4.3 4.4 4.5 Helbig, A. J., Seibold, I., Kocum, A., Liebers, D., Irwin, J., Bergmanis, U., Meyburg, B.-U., Scheller, W., Stubbe, M. & Bensch, S. (2005). Genetic differentiation and hybridization between greater and lesser spotted eagles (Accipitriformes: Aquila clanga, A. pomarina). The Journal of Ornithology, 146(3), 226-234.
- ^ 5.00 5.01 5.02 5.03 5.04 5.05 5.06 5.07 5.08 5.09 5.10 5.11 5.12 5.13 5.14 5.15 5.16 5.17 5.18 5.19 5.20 5.21 5.22 5.23 5.24 5.25 5.26 5.27 5.28 5.29 5.30 5.31 5.32 5.33 5.34 5.35 5.36 5.37 5.38 5.39 5.40 5.41 5.42 5.43 5.44 5.45 5.46 5.47 5.48 5.49 5.50 5.51 5.52 5.53 5.54 5.55 5.56 5.57 5.58 5.59 5.60 5.61 5.62 5.63 5.64 5.65 5.66 5.67 5.68 5.69 5.70 5.71 5.72 5.73 5.74 5.75 5.76 5.77 5.78 5.79 5.80 5.81 5.82 5.83 5.84 5.85 5.86 Ferguson-Lees, J.; Christie, D. Raptors of the World. Houghton Mifflin Harcourt. 2001. ISBN 0-618-12762-3.
- ^ 6.00 6.01 6.02 6.03 6.04 6.05 6.06 6.07 6.08 6.09 6.10 6.11 6.12 6.13 6.14 6.15 6.16 6.17 6.18 6.19 6.20 6.21 6.22 6.23 6.24 6.25 6.26 6.27 6.28 6.29 6.30 6.31 6.32 6.33 6.34 Naoroji, R., & Schmitt, N. J. (2007). Birds of prey of the Indian subcontinent. Om Books International.
- ^ 7.00 7.01 7.02 7.03 7.04 7.05 7.06 7.07 7.08 7.09 7.10 7.11 7.12 7.13 Karyakin, I. V. (2008). The Greater Spotted Eagle in the Volga region, Ural Mountains and western Siberia. Raptors Conservation, 11, 23-69.
- ^ 8.00 8.01 8.02 8.03 8.04 8.05 8.06 8.07 8.08 8.09 8.10 8.11 8.12 8.13 8.14 8.15 8.16 8.17 8.18 8.19 8.20 8.21 8.22 8.23 8.24 8.25 8.26 8.27 8.28 8.29 8.30 8.31 8.32 Brown, L. & Amadon, D. (1986) Eagles, Hawks and Falcons of the World. The Wellfleet Press. ISBN 978-1555214722.
- ^ 9.0 9.1 Väli, Ü. (2004). The greater spotted eagle Aquila clanga and the lesser spotted eagle A. pomarina: taxonomy, phylogeography and ecology. Tartu University Press.
- ^ 10.0 10.1 10.2 10.3 Watson, Jeff. The Golden Eagle. A&C Black. 2010. ISBN 978-1-4081-1420-9.
- ^ 11.0 11.1 11.2 11.3 11.4 11.5 Väli, Ü., Dombrovski, V., Treinys, R., Bergmanis, U., Daroczi, S. J., Dravecky, M., Ivanovski, V., Lontkowski, J, Maciorowski, G., Meyburg, B.-U., Mizera, T., Zeitz, R. & Ellegren, H. (2010). Widespread hybridization between the greater spotted eagle Aquila clanga and the lesser spotted eagle Aquila pomarina (Aves: Accipitriformes) in Europe. Biological Journal of the Linnean Society, 100(3), 725-736.
- ^ 12.0 12.1 Väli, Ü. (2015). Monitoring of spotted eagles in Estonia in 1994-2014: Stability of the lesser spotted eagle (Aquila pomarina) and decline of the greater spotted eagle (A. clanga). Slovak Raptor Journal, 9(1), 55.
- ^ Gill, F., Donsker, D., & Rasmussen, P. (Eds.). (2021). IOC world bird list. IOC.
- ^ Parry, S.J.; Clark, W.S.; Prakash, V. On the taxonomic status of the Indian Spotted Eagle Aquila hastata. Ibis. 2002, 144 (4): 665–675. doi:10.1046/j.1474-919X.2002.00109.x.
- ^ 15.0 15.1 15.2 Rasmussen, Pamela C.; Anderton, John C. Birds of South Asia - The Ripley Guide. Barcelona: Lynx Edicions. 2005. ISBN 84-87334-67-9.
- ^ 16.0 16.1 Väli, Ülo. Mitochondrial DNA sequences support species status for the Indian Spotted Eagle Aquila hastata. Bulletin of the British Ornithologists' Club. 2006, 126 (3): 238–242.
- ^ Wink, M., & Sauer-Gürth, H. (2004). Phylogenetic relationships in diurnal raptors based on nucleotide sequences of mitochondrial and nuclear marker genes. Raptors worldwide, 483-498.
- ^ 18.0 18.1 18.2 Lerner, H. R., & Mindell, D. P. (2005). Phylogeny of eagles, Old World vultures, and other Accipitridae based on nuclear and mitochondrial DNA. Molecular phylogenetics and evolution, 37(2), 327-346.
- ^ 19.0 19.1 Väli, Ü. (2002). Mitochondrial pseudo‐control region in old world eagles (genus Aquila). Molecular Ecology, 11(10), 2189-2194.
- ^ Jobling, James A. The Helm Dictionary of Scientific Bird Names. London: Christopher Helm. 2010: 110. ISBN 978-1-4081-2501-4.
- ^ 21.00 21.01 21.02 21.03 21.04 21.05 21.06 21.07 21.08 21.09 21.10 21.11 21.12 21.13 21.14 Global Raptor Information Network. 2021. Species account: Greater Spotted Eagle Clanga clanga. Downloaded from http://www.globalraptors.org on 18 Oct. 2021
- ^ Helbig, A. J., Seibold, I., Kocum, A., Liebers, D., Irwin, J., Bergmanis, U., Meyburg, B.-U., Scheller, W., Stubbe, M. & Bensch, S. (2005). Genetic differentiation and hybridization between greater and lesser spotted eagles (Accipitriformes: Aquila clanga, A. pomarina). journal of Ornithology, 146(3), 226-234.
- ^ Lerner, H., Christidis, L., Gamauf, A., Griffiths, C., Haring, E., Huddleston, C.J., Kabra, S., Kocum, A., Krosby, M., Kvaloy, K., Mindell, D., Rasmussen, P., Rov, N., Wadleigh, R., Wink, M. & Gjershaug, J.O. (2017). Phylogeny and new taxonomy of the Booted Eagles (Accipitriformes: Aquilinae). Zootaxa, 4216(4), 301-320.
- ^ 24.0 24.1 24.2 24.3 24.4 24.5 24.6 Lontkowski, J., & Maciorowski, G. (2010). Identification of juvenile greater spotted eagle, lesser spotted eagle and hybrids. Dutch Birding, 32, 384-397.
- ^ 25.0 25.1 25.2 25.3 25.4 25.5 25.6 Cramp, S., & Simmons, K. E. L. (1980). Vol. II: Hawks to bustards. Oxford [etc.]: Oxford University Press.
- ^ Brown, L. (1977). Eagles of the World. Universe Books.
- ^ Christensen, S., Nielsen, B. P., Porter, R. F., & Willis, I. (1974). Flight identification of European raptors. British Birds, 64(6), 247-266.
- ^ Dombrovski, V. C., & Demongin, L. (2006). On geographic variability of some diagnostic characteristics of the greater spotted eagle (Aquila clanga). Á Ornithologia, 33.
- ^ 29.0 29.1 29.2 29.3 CRC Handbook of Avian Body Masses, 2nd Edition by John B. Dunning Jr. (Editor). CRC Press (2008), ISBN 978-1-4200-6444-5.
- ^ 30.00 30.01 30.02 30.03 30.04 30.05 30.06 30.07 30.08 30.09 30.10 30.11 30.12 30.13 30.14 30.15 30.16 30.17 30.18 30.19 30.20 30.21 30.22 30.23 30.24 30.25 30.26 Meyburg, B.-U., G. M. Kirwan, and E. F. J. Garcia (2020). Greater Spotted Eagle (Clanga clanga), version 1.0. In Birds of the World (J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.grseag1.01
- ^ Jeyarajasingam, A. (2012). A Field Guide to the birds of Peninsular Malaysia and Singapore. Oxford University Press.
- ^ Freedman, B., & Keith, T. L. (Eds.). (1998). Encyclopedia of Endangered Species (Vol. 2). Gale/Cengage Learning.
- ^ Cieslak, M., & Dul, B. (2006). Feathers: Identification for Bird Conservation. Warsaw: Natura Publishing House.
- ^ 34.0 34.1 34.2 34.3 Dementiev, G. P., Gladkov, N. A., Ptushenko, E. S., Spangenberg, E. P., & Sudilovskaya, A. M. (1966). Birds of the Soviet Union, vol. 1. Israel Program for Scientific Translations, Jerusalem.
- ^ 35.0 35.1 Parry, S. J. (2001). The booted eagles (Aves: Accipitridae): perspectives in evolutionary biology. University of London, University College London (United Kingdom).
- ^ Dombrovski, V. C. (2006). Morphological Characteristics and Diagnostic Features of the Greater Spotted (Aquila clanga), Lesser Spotted (A. pomarina) eagles, and their hybrids. Á Ornithologia, 33.
- ^ Dresser, H. A. (1902). A Manual of Palæarctic Birds, Vol. 1.
- ^ Hartert, E. (1912). Die Vögel der paläarktischen Fauna: systematische Übersicht der in Europa, Nord-Asien und der Mittelmeerregion vorkommenden Vögel (Vol. 2). Friedlaender & Sohn.
- ^ Roberts, T. J. (1991). The Birds of Pakistan: Passeriformes: Pittas to Buntings (Vol. 2). Oxford University Press.
- ^ Forsman, D. (1999). The Raptors of Europe and the Middle East: a Handbook of Field Identification. London: T & AD Poyser.
- ^ Clark, W.S. (1990). Spotted Eagle with rufous nape patch. British Birds. 83(9): 397-399.
- ^ 42.0 42.1 Bergmanis, U. (1996). On the taxonomy of the lesser spotted eagle Aquila pomarina and greater spotted eagle A. clanga. Eagle studies, 199-207.
- ^ Svensson, Lars. Underwing pattern of Steppe, Spotted and Lesser Spotted Eagles. International Bird Identification: Proceedings of the 4th International Identification Meeting. Eilat: International Birdwatching Centre Eilat: 12–14. 1–8 November 1986.
- ^ Corso, A. (1999). Separating juvenile Imperial and Greater Spotted Eagles, in particular of pale morph ‘fulvescens’. Dutch Birding. 21(3): 150-151.
- ^ Golemanski, Vasil and collective. Red Book of Bulgaria, Volume II. Bulgarian Academy of Sciences. 2015: 77. ISBN 978-954-9746-18-1.
- ^ Gabor, M. (1998). Annotated List of the Birds of Hungary.
- ^ Rokkanen, S. (2019). The expansion of protected area network and the conservation status change of breeding birds in Finland 1996-2015.
- ^ 48.0 48.1 48.2 Flint, V. E. (1984). A field guide to birds of the USSR: including Eastern Europe and Central Asia. Princeton University Press.
- ^ 49.0 49.1 Wassink, A., & Oreel, G. J. (2007). The Birds of Kazakhstan. A. Wassink.
- ^ Dharmakuarsinhji, K.S. (1955). Birds of Saurashtra. Dil Bahar.
- ^ Prakash, V. (1988). Greater Spotted Eagle (Aquila clanga) breeding in Keoladeo National Park, Bharatpur. J Bombay Nat Hist Soc, 85(2), 418.
- ^ Swinhoe, L. C. C., & Barnes, L. H. (1885). On the Birds of Central India.–Part II. Ibis, 27(2), 124-138.
- ^ Ramadan-Jaradi, G., & Ramadan-Jaradi, M. (1999). An Updated Checklist of the Birds of Lebanon. Sandgrouse, 21, 132-170.
- ^ Nankinov, D. (1992). Check list of Bbird Species and subspecies in Bulgaria. Avocetta, 16(1), 1.
- ^ Bijlsma, R. G. (2001). Common and Scarce Birds of the Netherlands. GMB Uitgeverij.
- ^ de Bruin, S., van Dijk, A. and Koops, E. (1998). Bastaardarend in Lauwersmeer in augustus 1997. Dutch Birding. 20(6): 283-285.
- ^ Brown, A., & Grice, P. (2010). Birds in England. A&C Black.
- ^ 58.0 58.1 58.2 Kren, J. (2000). Birds of the Czech Republic. Helm.
- ^ Töpfer, Till. Nachweise seltener Vogeltaxa (Aves) in Sachsen aus der ornithologischen Sammlung des Museums für Tierkunde Dresden [Records of rare bird taxa (Aves) in Saxony from the ornithological collection of the Zoological Museum Dresden]. Faunistische Abhandlungen. 2007, 26 (3): 63–101 (德语及英语).
- ^ Thévenot, M., R. Vernon, and P. Bergier (2003). The Birds of Morocco: An Annotated Check-list. British Ornithologists' Union Check-list 20. British Ornithologists' Union, Tring, United Kingdom.
- ^ Zimmerman, D. A., D. A. Turner, and D. J. Pearson (1996). Birds of Kenya and Northern Tanzania. Princeton University Press, Princeton, NJ, USA.
- ^ Leonard, P.M. (1998). New to Zambia: Greater Spotted Eagle Aquila clanga. Zambia Bird Report. 1997: 3-5.
- ^ Vallotton, L. (2008). Possible first record of a Greater Spotted Eagle Aquila clanga in Botswana. Gabar, 22: 28-30.
- ^ Argandeval, M. E. (1983). Raspredelenie i chislennost khishchnykh ptits v gornykh landshaftakh tsentralnogo i vostochnogo Afganistana [Distribution and number of birds of prey in mountain landscapes of the Central and Eastern Afghanistan]. In Ekologiya khishchnykh ptits [Ecology of birds of prey]. Materials of the 1st Meeting on Ecology and Conservation of Birds of Prey, Moscow (pp. 16-18).
- ^ Spierenburg, P. (2005). Birds in Bhutan. Status and distribution, 1-383.
- ^ Verheugt, W. J. M., H. Skov, and F. Danielsen (1993). Notes on the birds of the tidal lowlands and floodplains of South Sumatra Province, Indonesia. Kukila 6(2):53–84.
- ^ Kirwan, G. M. & Yousef, R. (2002). Raptor migration in Israel and the Middle East: A summary of 30 years of field research. International Birding & Research Center in Eilat.
- ^ 68.0 68.1 68.2 Maciorowski, G., Galanaki, A., Kominos, T., Dretakis, M., & Mirski, P. (2019). The importance of wetlands for the Greater Spotted Eagle Clanga clanga wintering in the Mediterranean Basin. Bird Conservation International, 29(1), 115-123.
- ^ de Juana, E., and E. Garcia (2015). The Birds of the Iberian Peninsula. Bloomsbury, London, United Kingdom.
- ^ Dubois, P. J., P. Le Maréchal, G. Olioso & P. Yésou (2008). Nouvel Inventaire des Oiseaux de France. Delachaux et Niestlé, Paris, France.
- ^ Handrinos, G., & Akriotis, T. (1997). The Birds of Greece. Christopher Helm.
- ^ Handrinos, G. (1987). The significance of Greece for wintering and migrating raptors. Ricerche Biologia Selvaggina, 12, 99-113.
- ^ 73.0 73.1 Ash, C. P., & Atkins, J. D. (2009). Birds of Ethiopia and Eritrea: an Atlas of Distribution. A&C Black.
- ^ Murdoch, D. A., & Betton, K. F. (2008). A Checklist of the Birds of Syria. Ornithological Society of the Middle East, Caucasus and Central Asia.
- ^ 75.0 75.1 Shirihai, H., Dovrat, E., Christie, D. A., & Harris, A. (1996). The Birds of Israel (Vol. 876). London: Academic Press.
- ^ Kirwan, G. M., Martins, R. P., Eken, G., & Davidson, P. (1999). A Checklist of the Birds of Turkey. Sandgrouse, 20, 1-32.
- ^ Patrikeev, M., & Harper, G. H. (2004). Birds of Azerbaijan. Pensoft.
- ^ Scott, D. A., & Adhami, A. (2006). An updated Checklist of the Birds of Iran. Podoces, 1(1/2), 1-16.
- ^ Ali, S., & Ripley, S. D. (1980). Handbook of the birds of India and Pakistan, together with those of Bangladesh, Nepal, Bhutan, and Sri Lanka. Oxford University Press.
- ^ Jerdon, T. C. (1862). The Birds of India: Being a Natural History of All the Birds Known to Inhabit Continental India, with Descriptions of the Species, Genera, Families, Tribes, and Orders, and a Brief Notice of Such Families as are Not Found in India, Making it a Manual of Ornithology Specially Adapted for India (Vol. 1). author.
- ^ Thewlis, R. M., R. J. Timmins, T. D. Evans, and J. W. Duckworth (1998). The Conservation Status of Birds in Laos: a Review of Key Species. Bird Conservation International 8(Supplement):1–159.
- ^ 82.0 82.1 82.2 82.3 Mallalieu, M. (2007). Greater spotted eagles Aquila clanga in central Thailand. Forktail, 23.
- ^ Duckworth, J.W. and Tizard, R.J. (2003). W.W. Thomas’s bird records from Laos, principally Vientiane, 1966–1968 and 1981–1983. Forktail 19:63–84.
- ^ 84.0 84.1 84.2 84.3 Wells, D. R. (1999). The Birds of the Thai-Malay Peninsula. London: Academic Press.
- ^ Brazil, M. (2009). Birds of East Asia: China, Taiwan, Korea, Japan, and Russia. A&C Black.
- ^ Gore, M. E. J., & Won, P. O. (1971). The Birds of Korea Royal Asiatic Society. Korean Branch, Seoul.
- ^ 87.0 87.1 87.2 Maciorowski, G., Mirski, P., Kardel, I., Stelmaszczyk, M., Mirosław-Swiątek, D., Chormański, J. and Okruszko, T. (2015). Water regime as a key factor differentiating habitats of spotted eagles Aquila clanga and Aquila pomarina in Biebrza Valley (NE Poland). Bird Study. 62(1): 120–125.
- ^ Smith, K. D. (1957). An annotated check list of the birds of Eritrea. Ibis, 99(2), 307-337.
- ^ Nikolaus, G. (1987). Distribution atlas of Sudan's birds with notes on habitat and status. Zoologisches Forschungsinstitut und Museum Alexander Koenig.
- ^ 90.0 90.1 Esra, P., Ulusoy, E., & Vural, D. (2018). An Unusual Record of Greater Spotted Eagle (Clanga clanga) in Winter in Ankara, Turkey. Commagene Journal of Biology, 2(1), 30-33.
- ^ Khan, M.M.H. (2005). Species diversity, relative abundance and habitat use of the birds in the Sundarbans East Wildlife Sanctuary, Bangladesh. Forktail. 21: 79–86.
- ^ 92.0 92.1 Lobley, G.R. (2007). Wintering of Greater Spotted Eagle Aquila clanga and Eastern Imperial Eagle A. heliaca in the Arabian Peninsula. Sandgrouse. 29(2): 177- 182.
- ^ Hamidi, N., & Musavi, S. B. (2006). Status of Imperial Eagle Aquila heliaca and Greater Spotted Eagle Aquila clanga in Hormozgan Province, southern Iran. Podoces, 1(1-2), 67-70.
- ^ Salim, M. A., Abed, S. A., & Porter, R. F. (2021). The ornithological importance of the southern marshes of Iraq. In Southern Iraq's Marshes (pp. 351-375). Springer, Cham.
- ^ 95.0 95.1 95.2 Väli, Ü., Dombrovski, V., & Mirski, P. (2021). Greater spotted eagle Clanga clanga. In Migration Strategies of Birds of Prey in Western Palearctic (pp. 88-100). CRC Press.
- ^ 96.0 96.1 96.2 Shirihai, H., R. Yosef, D. Alon, G. M. Kirwan, and R. Spaar (2000). Raptor migration in Israel and the Middle East: a summary of 397 years of field research. International Birding & Research Center in Eilat, Israel.
- ^ 97.0 97.1 Üner, Ö., Boyla, K. A., Bacak, E., Birel, E., Çelikoba, İ., Dalyan, C., Tabur, E. and Yardim, Ü. (2010). Spring migration of soaring birds over the Bosphorus, Turkey. Sandgrouse. 32(1): 20-33.
- ^ 98.0 98.1 Babbington, J. & Roberts, P. (2012). Greater Spotted Eagles Aquila clanga summering in Saudi Arabia? Sandgrouse. 34(1): 65-66.
- ^ Bishop, K.D. (1999). Preliminary notes on some birds in Bhutan. Forktail. 15: 87–91.
- ^ Tordoff, A.W. (2002). Raptor migration at Hoang Lien Nature Reserve, northern Vietnam. Forktail. 18: 45–48.
- ^ DeCandido, R., Nualsri, C., Allen, D. and Bildstein, K.L. (2004). Autumn raptor migration at Chumphon, Thailand: a globally significant raptor migration watch site. Forktail. 20: 49–54.
- ^ de Bont, M. (2009). Bird observations from south-east Sudan. Bull. Afr. Bird Club 16(1): 37-52.
- ^ Hedenström, A. (1997). Predicted and observed migration speed in Lesser Spotted Eagle Aquila pomarina. Ardea. 85(1): 29-36.
- ^ 104.0 104.1 Demey, R., Lack, P. & Webb, R. (2007). Africa Round-up. Satellite-tracked Greater Spotted Eagles. Bull. African Bird Club. 14(1): 14-15.
- ^ 105.0 105.1 105.2 Alexeyenko, M. N. & Fefelov, I. V. (2008). Autumn Migration of Greater Spotted Eagle in the South Baikal Migratory Pass. Study and protection of great and lesser spotted eagles in Severnaya , 26.
- ^ Sheng, T.A.W. (2012). Hunting strategy of Greater Spotted Eagle Aquila clanga at Permatang Pauh, Pulau Pinang. Suara Enggang. 20(2): 8.
- ^ Strick, J., Vercammen, P., Judas, J. and Combreau, O. (2011). Satellite tracking of a rehabilitated Greater Spotted Eagle Aquila clanga. Zoology in the Middle East. 54(Suppl. 3): 103-106.
- ^ Meyburg, B.-U., Eichacker, X., Meyburg, C. & Paillat, P. (1995). Migrations of an adult Spotted Eagle tracked by satellite. British Birds. 88(8): 357-361.
- ^ Pérez-García, J. M., Sellis, U., & Väli, Ü. (2014). Winter ranging behaviour of a greater spotted eagle (Aquila clanga) in southeast Spain during four consecutive years. Slovak Raptor Journal, 8(2), 123.
- ^ Dowsett, R.J., Aspinwall, D.R. & Leonard, P.M. (1999). Further additions to the avifauna of Zambia. Bull. Brit. Orn. Club. 119(2): 94–103.
- ^ 111.00 111.01 111.02 111.03 111.04 111.05 111.06 111.07 111.08 111.09 Dombrovski, V. (2010). The diet of the greater spotted eagle (Aquila clanga) in Belarusian Polesie. Slovak Raptor Journal, 4, 23–36.
- ^ Marti, C. D., Korpimäki, E., & Jaksić, F. M. (1993). Trophic structure of raptor communities: a three-continent comparison and synthesis. In Current ornithology (pp. 47-137). Springer, Boston, MA.
- ^ 113.0 113.1 113.2 113.3 113.4 Graszynski, K., Komischke, B., & Meyburg, B. U. (2002). On the biology of the greater spotted eagle (Aquila clanga Pallas 1811). Raptors in the New Millennium (ed. Yosef, R., Miller, ML & Pepler, D.), 62-75.
- ^ Martínez, J. E., Zuberogoitia, I., Gómez, G., Escarabajal, J. M., Cerezo, E., Jiménez-Franco, M. V., & Calvo, J. F. (2014). Attack success in Bonelli's Eagle Aquila fasciata. Ornis Fennica, 91(2), 67.
- ^ 115.0 115.1 115.2 Väli, Ü. & Lõhmus, A. (2002). Parental care, nestling growth and diet in a Spotted Eagle Aquila clanga nest. Bird Study. 49(1): 93-95.
- ^ Ivanovsky, V. V. (2012). Birds of Prey of the Belarusian Poozerie Monograph. Voronezh State University.
- ^ Alivizatos, H., Papandropoulos, D., & Zogaris, S. (2004). Winter diet of the Greater Spotted Eagle (Aquila clanga) in the Amvrakikos wetlands, Greece. Journal of Raptor Research, 38(4), 371-374.
- ^ 118.0 118.1 118.2 118.3 Pérez-García, J. M., Marco-Tresserras, J., & Orihuela-Torres, A. (2020). Winter diet and lead poisoning risk of Greater Spotted Eagles Clanga clanga in southeast Spain. Bird Study, 67(2), 224-231.
- ^ Pukinsky, Y. B. (2015). To the diet of the great spotted eagle Aquila clanga in the Leningrad region during the nesting period. Russian Ornithological Journal, 1201: 3697-3698.
- ^ Georgievich, V. (2018). About the diet of the great spotted eagle Aquila clanga at the mouth of the Belaya River.
- ^ 121.0 121.1 121.2 121.3 121.4 121.5 Karyakin, I. V. (2008). The Greater Spotted Eagle in the Altai-Sayan region.
- ^ 122.0 122.1 122.2 122.3 Prakash, V. (1988). The general ecology of raptors in Keoladeo National Park, Bharatpur (Doctoral dissertation, Ph. D. thesis. Bombay University, Mumbai, India).
- ^ 123.0 123.1 123.2 Naoroji, R. (1990). Predation by Aquila eagles on nestling storks and herons in Keoladeo National Park, Bharatpur. Journal of the Bombay Natural History Society, 80087(1), 37-46.
- ^ Selover, W. C. (2004). The Birds of Bharatpur. Indian Birds.
- ^ Urfi, A. J. (2011). The Painted Stork: ecology and conservation. Springer Science & Business Media.
- ^ 126.0 126.1 Zacharias, V. J., Leisler, B., & Leisler, A. (2019). Greater Spotted Eagle Clanga clanga Pallas at Vembanadu lake, Kerala, South India. Indian Forester, 145(10), 1007-1008.
- ^ 127.0 127.1 Alivizatos, H., & Goutner, V. (2021). Diet composition, guild structure and trophic relationships of wintering birds of prey in an estuarine wetland (The Evros Delta National Park, Greece). Ecologica Montenegrina, 39, 15-29.
- ^ Levy, N. and Yom-Tov, Y. (1991). Activity and status of Cranes Grus grus wintering in Israel. Sandgrouse. 13(2): 58–72.
- ^ Johnsgard, P. A. (1983). Cranes of the World: Japanese Crane (Grus japonensis). Cranes of the World, by Paul Johnsgard, 21.
- ^ 130.0 130.1 Maciorowski, G., Lontkowski, J., & Mizera, T. (2014). The spotted eagle–vanishing bird of the marshes. Agencja Promocyjno-Wydawnicza UNIGRAF, Bydgoszcz, 308.
- ^ Ivanovsky, Vladimir V. "Trophic Links Between Birds of Prey and Their Preys and Their Competitors in Wetlands During Nesting Period." Raptors Conservation 38 (2019).
- ^ Meyburg, B.-U., P. F. D. Boesman, J. S. Marks, and G. M. Kirwan (2020). Lesser Spotted Eagle (Clanga pomarina), version 1.0. In Birds of the World (J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA.
- ^ Demerdzhiev, D. A., Popgeorgiev, G. S., Dobrev, D. D., Arkumarev, V. S., & Terziev, N. G. (2019). Habitat requirements of the Lesser Spotted Eagle Clanga pomarina Brehm, 1831 (Aves: Accipitridae) at the southern periphery of the distribution range (Southeast Bulgaria). Acta Zoologica Bulgarica, Supplement, 14, 35-65.
- ^ Al-Sheikhly, O. F., Al-Barazangi, A. N., Haba, M. K., Fazaa, N. A., Abdulzahra, H. K., Turab, M. K. A., & Al-Azawi, A. J. (2017). Ring Recoveries from Steppe Eagles and Eastern Imperial Eagles from the Russian and Kazakhstan Breeding Populations and a Review of Major Threats to Eagles in Iraq. Raptors Conservation, 35.
- ^ Nikolenko, E. G. (2013). The Project" Eagles of Russia": Results of Eagle Ringing in 2013. Raptors Conservation, (27).
- ^ Porter, R., & Beaman, M. (1985). A resume of raptor migration in Europe and the Middle East. Conservation Studies on Raptors. ICBP Technical Publication, 5, 237-242.
- ^ Horváth, M., Solti, B., Fatér, I., Juhász, T., Haraszthy, L., Szitta, T., Bállok, Z. & Pásztory-Kovács, S. (2018). Temporal changes in the diet composition of the Eastern Imperial Eagle (Aquila heliaca) in Hungary. Ornis Hungarica, 26(1), 1-26.
- ^ Katzner, T. E., Bragin, E. A., Knick, S. T., & Smith, A. T. (2006). Spatial structure in the diet of imperial eagles Aquila heliaca in Kazakhstan. Journal of Avian Biology, 37(6), 594-600.
- ^ Karyakin I.V. (2015). The Steppe Eagle (Aquila nipalensis). – Russian Raptor Research and Conservation Network.
- ^ Karyakin, I.V.,Nikolenko, E. G., Zinevich, L. S. & Pulikova, G. I. (2017). Steppe Eagle in the Karaganda Region, Kazakhstan. Raptors Conservation, 35.
- ^ Bekmansurov, R. H., Karyakin, I. V., & Shnayder, E. P. (2015). On Eastern Imperial Eagle (Aquila heliaca) breeding in atypical habitat under competitive conditions with other eagle species. Slovak Raptor Journal, 9(1), 95.
- ^ Bishop, K. David. Preliminary notes on some birds in Bhutan (PDF). Forktail. 1999, 15: 87–91 [18 July 2014]. (原始内容 (PDF)存档于26 July 2014).
- ^ Sashikumar, C. (2004). Aquila eagles in Kerala, India. News Ornis, 1(4), 53-54.
- ^ Domashevsky, S. Franchuk, M. Komarnitsky, I. & Chovan, A. (2015). Expedition to study the Great Spotted Eagle in the north-west of Ukraine in 2015. Ukrainian Raptor Center.
- ^ Meyburg, B.U., Mizera, T., Maciorowski, G., Dylawerski, M. & Smyk, A. (1995). Juvenile Spotted Eagle apparently killed by Eagle Owl. British Birds. 88(8): 376.
- ^ Penteriani, V., & del Mar Delgado, M. (2019). The eagle owl. Bloomsbury Publishing.
- ^ 147.0 147.1 147.2 Karyakin, I. V. & Levin, A. S. (2008). The Greater Spotted Eagle in Kazakhstan. — Research and Conservation of the Spotted Eagles.
- ^ 148.0 148.1 148.2 148.3 148.4 Shukov, P. M., Bakka, S. V., & Kiseleva, N. Y. (2021). The Greater and Lesser Spotted Eagles in the Center of European Russia. In IOP Conference Series: Earth and Environmental Science (Vol. 723, No. 2, p. 022096). IOP Publishing.
- ^ Karyakin, I. V. (2008). Ecology of the Greater Spotted Eagle in Volga-Ural Region. — Research and Conservation of the Spotted Eagles.
- ^ Collar, N. J., A. V. Andreev, S. Chan, M. J. Crosby, S. Subramanya, & J. A. Tobias, Editors (2001). Threatened Birds of Asia: the BirdLife International Red Data Book. BirdLife International, Cambridge, United Kingdom.
- ^ Harrison, C. J. O. & P. Castell (2002). Bird Nests, Eggs and Nestlings of Britain and Europe with North Africa and the Middle East. Second revised edition. HarperCollins, London, United Kingdom.
- ^ 152.0 152.1 152.2 Dombrovski, V. C. (2019). Timing, Diet and Parental Care in a Spotted Eagle Nest in Chernobyl Exclusion Zone (Belarus) in 2018 as Revealed by Camera Trap. Raptors Conservation, 38.
- ^ Karyakin, I., Nikolenko, E., & Bekmansurov, R. (2009). Results of Monitoring of Greater Spotted Eagle and Imperial Eagle Breeding Groups in the Altai Pine Forests in 2009, Russia. Raptors Conservation, (17).
- ^ Simmons, R. (1988). Offspring quality and the evolution of cainism. Ibis, 130(3), 339-357.
- ^ Stinson, C. H. (1979). On the selective advantage of fratricide in raptors. Evolution, 1219-1225.
- ^ Meyburg, B. U. (2002). On Cainism in the lesser spotted eagle (Aquila pomarina) and a possible explanation for the phenomenon in this and other eagle species. Raptors in the New Millennium, 53-61.
- ^ 157.0 157.1 Meyburg, B. U., & Pielowski, Z. (1991). Cainism in the Greater Spotted Eagle Aquila clanga. Birds of Prey Bull, 4, 143-148.
- ^ Karyakin, I. V. (2008). Ecology of the Greater Spotted Eagle in Western Siberia. — Research and Conservation of the Spotted Eagles.
- ^ 159.0 159.1 Meyburg, B., Meyburg, C., Mizera, T., Maciorowski, G., & Kowalski, J. (2005). Family break up, departure, and autumn migration in Europe of a family of Greater Spotted Eagles (Aquila clanga) as reported by satellite telemetry. Journal of Raptor Research, 39(4), 462.
- ^ Lõhmus, A., & Väli, Ü. (2001). Interbreeding of the greater Aquila clanga and lesser spotted eagle A. pomarina. Acta ornithoecol, 4, 377-384.
- ^ Meyburg, B. U., & Meyburg, C. (2007). Post-fledging behavior and outward migration of a hybrid Greater× Lesser Spotted Eagle (Aquila clanga× A. pomarina). Journal of Raptor Research, 41(2), 165-170.
- ^ Väli, Ülo; Lõhmus, Asko. Nestling characteristics and identification of the lesser spotted eagle Aquila pomarina, greater spotted eagle A. clanga, and their hybrids. Journal of Ornithology. 2004, 145 (3): 256–263. S2CID 20893726. doi:10.1007/s10336-004-0028-7.
- ^ 163.0 163.1 Treinys, R. (2005). The Greater Spotted Eagle (Aquila clanga): previous, current status and hybridisation in Lithuania. Acta zoologica lituanica, 15(1), 31-38.
- ^ 164.0 164.1 Maciorowski, G., Mirski, P., & Väli, Ü. (2015). Hybridisation dynamics between the Greater Spotted Eagles Aquila clanga and Lesser Spotted Eagles Aquila pomarina in the Biebrza River Valley (NE Poland). Acta Ornithologica, 50(1), 33-41.
- ^ Maciorowski, G., & Mirski, P. (2014). Habitat alteration enables hybridisation between lesser spotted and greater spotted eagles in north-east Poland. Bird Conservation International, 24(2), 152-161.
- ^ Dravecký, M., Sellis, U., Bergmanis, U., Dombrovski, V., Lontkowski, J., Maciorowski, G., Maderic, B., Meyburg, B. U., Mizera, T., Stoj, M., Treinys, R. & Wójciak, J. (2008). Colour ringing of the Spotted Eagles (and their hybrids) in Europe-a review. Raptor Journal, 2(2008), 37-52.
- ^ Gorban, I. (1996). Lesser and Greater Spotted Eagle Aquila pomarina and A. clanga in Ukraine. Eagle Studies. World Working Group on Birds of Prey, Berlin, London & Paris, 301-302.
- ^ Volke, V. (1996). The status of the Greater Spotted Eagle Aquila clanga and Lesser Spotted Eagle A. pomarina in Estonia. Eagle studies, 285-289.
- ^ Kalyakin, M. V., & Voltzit, O. V. (2006). The Atlas of the Birds of Moscow City Project: history, methods and first results. Bird Census, 63.
- ^ Hussain, M.M., A. Doley, R. Dutta & H. Singha (2019). Sighting of Greater Spotted Eagle Clanga clanga in Assam University, Silchar Campus with its current distribution in Assam, India. Bird-osoar #34, In: Zoo’s Print 34(9): 17–26.
- ^ Santhakumar, B., Ali, A. M. S., & Arun, P. R. (2016). Status of Greater Spotted Eagle Clanga clanga in Tamil Nadu and Puducherry, India. Indian Birds, 11, 71-74.
- ^ Kataria, A. K., Kataria, N., & Kumawat, R. N. (2016). Effect of environmental elements on migration pattern of eagles at Jorbeer conservation reserve, Bikaner, Rajasthan, India. Research Journal of Life Sciences, Bioinformatics, Pharmaceuticals and Chemical sciences, 2016c, 2(3), 90-101.
- ^ Ananian, V. (2008). On the finds of Greater Spotted Eagle in Armenia. Изучение и охрана большого и малого подорликов в Северной, 34.
- ^ 174.0 174.1 Väli, Ülo; Lõhmus, Asko. Suur-konnakotkas ja tema kaitse Eestis [The Greater Spotted Eagle and its conservation in Estonia]. Hirundo. 2000,. Supplement 3: 1–50.
- ^ Khaleghizadeh, A. (2004). Notes on three rare raptors in the Anzali wetlands, Iran. Sandgrouse. 26(2): 155.
- ^ Meadows, B.S. (2011). A note on occurrence at man-made habitats of wintering Greater Spotted Aquila clanga and Eastern Imperial Eagles A. heliaca in the coastal belt of eastern Saudi Arabia. Sandgrouse. 33(2): 98-101.
- ^ Dixon, A., Maming, R., Gunga, A., Purev-Ochir, G., & Batbayar, N. (2013). The problem of raptor electrocution in Asia: case studies from Mongolia and China. Bird Conservation International, 23(4), 520-529.
- ^ Quigxia, Z. (1996). Winter ecology of Aquila clanga in Lishan Nature Reserve. Sichuan Journal of Zoology 15: 170-172.
- ^ 179.0 179.1 Meyburg, B. U., Haraszthy, L., Strazds, M., & Schäffer, N. (2001). European species action plan for greater spotted eagle. European Union Action Plans for Eight Priority Birds Species. Europ Comm, Luxembourg.
- ^ Bakka, S. V., Kiseleva, N. Y., & Shukov, P. M. (2020). Installing of Artificial Nests as a Method of the Large Birds of Prey Population Management in the Center of European Russia: Successes, Problems, Prospects. In IOP Conference Series: Earth and Environmental Science (Vol. 543, No. 1, p. 012015). IOP Publishing.
- ^ Väli, Ü., Dombrovski, V., Dzmitranok, M., Maciorowski, G., & Meyburg, B. U. (2019). High genetic diversity and low differentiation retained in the European fragmented and declining Greater Spotted Eagle (Clanga clanga) population. Scientific reports, 9(1), 1-11.
- ^ Väli, Ü., Treinys, R., & Poirazidis, K. (2004). Genetic structure of Greater Aquila clanga and Lesser Spotted Eagle A pomarina populations: implications for phylogeography and conservation. Raptors Worldwide. World Working Group on Birds of Prey & BirdLife Hungary, Budapest, 473-482.
| ||||||||||||
| 这是一篇鹰形目小作品。您可以通过编辑或修订扩充其内容。 |