
自噬:修订间差异
Sing20041130(留言 | 贡献) 內容擴充 |
Sing20041130(留言 | 贡献) 內容擴充 |
||
| 第1行: | 第1行: | ||
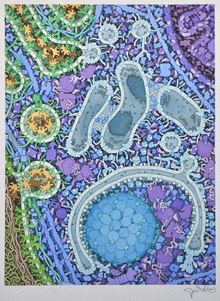
[[File:Autophagy diagram PLoS Biology.jpg|thumb|right|(A) 自噬示意圖; (B) 果蠅幼蟲的脂肪體自噬結構的電子顯微鏡照片; (C) 螢光標記的自噬體飢餓小鼠肝細胞]] |
[[File:Autophagy diagram PLoS Biology.jpg|thumb|right|(A) 自噬示意圖; (B) 果蠅幼蟲的脂肪體自噬結構的電子顯微鏡照片; (C) 螢光標記的自噬體飢餓小鼠肝細胞]] |
||
'''自噬'''({{lang-en|Autophagy}},或稱'''自體吞噬''')是一個涉及到[[細胞]]自身結構通過[[溶酶體]]機制,負責將受損的細胞器、錯誤折疊的蛋白及其他大分子物質等運送至溶酶體降解並再利用的進化保守過程。自噬是廣泛存在於真核細胞的現象,並且可分為巨自噬、微自噬和分子伴侶介導的自噬三大類。這是一個受到緊密調控的步驟,此步驟是細胞生長、[[細胞分化|發育]]與[[穩態]]中的常規步驟,幫助細胞產物在合成、降解以及接下來的循環中保持一個平衡狀態。目前已有多份研究表明自噬在許多細胞的分化進程中被不同程度地激活<ref>{{cite journal |last1=Guan |first1=JL |last2=Simon |first2=AK |last3=Prescott |first3=M |last4=Menendez |first4=JA |last5=Liu |first5=F |last6=Wang |first6=F |last7=Wang |first7=C |last8=Wolvetang |first8=E |last9=Vazquez-Martin |first9=A |last10=Zhang |first10=J |title=Autophagy in stem cells. |journal=Autophagy |date=2013-06-01 |volume=9 |issue=6 |pages=830-49 |doi=10.4161/auto.24132 |pmid=23486312 |url=https://pubmed.ncbi.nlm.nih.gov/23486312-autophagy-in-stem-cells/ |accessdate=2020-01-28}}</ref>,例如參與血管生成<ref>{{cite journal |last1=Torisu |first1=T |last2=Torisu |first2=K |last3=Lee |first3=IH |last4=Liu |first4=J |last5=Malide |first5=D |last6=Combs |first6=CA |last7=Wu |first7=XS |last8=Rovira |first8=II |last9=Fergusson |first9=MM |last10=Weigert |first10=R |last11=Connelly |first11=PS |last12=Daniels |first12=MP |last13=Komatsu |first13=M |last14=Cao |first14=L |last15=Finkel |first15=T |title=Autophagy regulates endothelial cell processing, maturation and secretion of von Willebrand factor. |journal=Nature medicine |date=2013-10 |volume=19 |issue=10 |pages=1281-7 |doi=10.1038/nm.3288 |pmid=24056772 |url=https://pubmed.ncbi.nlm.nih.gov/24056772-autophagy-regulates-endothelial-cell-processing-maturation-and-secretion-of-von-willebrand-factor/ |accessdate=2020-01-28}}</ref>、成骨分化<ref>{{cite journal |last1=Pantovic |first1=A |last2=Krstic |first2=A |last3=Janjetovic |first3=K |last4=Kocic |first4=J |last5=Harhaji-Trajkovic |first5=L |last6=Bugarski |first6=D |last7=Trajkovic |first7=V |title=Coordinated time-dependent modulation of AMPK/Akt/mTOR signaling and autophagy controls osteogenic differentiation of human mesenchymal stem cells. |journal=Bone |date=2013-01 |volume=52 |issue=1 |pages=524-31 |doi=10.1016/j.bone.2012.10.024 |pmid=23111315 |url=https://pubmed.ncbi.nlm.nih.gov/23111315-coordinated-time-dependent-modulation-of-ampkaktmtor-signaling-and-autophagy-controls-osteogenic-differentiation-of-human-mesenchymal-stem-cells/ |accessdate=2020-01-28}}</ref>、脂肪生成<ref>{{cite journal |last1=Nuschke |first1=A |last2=Rodrigues |first2=M |last3=Stolz |first3=DB |last4=Chu |first4=CT |last5=Griffith |first5=L |last6=Wells |first6=A |title=Human mesenchymal stem cells/multipotent stromal cells consume accumulated autophagosomes early in differentiation. |journal=Stem cell research & therapy |date=2014-12-17 |volume=5 |issue=6 |pages=140 |doi=10.1186/scrt530 |pmid=25523618 |url=https://pubmed.ncbi.nlm.nih.gov/25523618-human-mesenchymal-stem-cellsmultipotent-stromal-cells-consume-accumulated-autophagosomes-early-in-differentiation/ |accessdate=2020-01-28}}</ref>、神經發生<ref>{{cite journal |last1=Vázquez |first1=P |last2=Arroba |first2=AI |last3=Cecconi |first3=F |last4=de la Rosa |first4=EJ |last5=Boya |first5=P |last6=de Pablo |first6=F |title=Atg5 and Ambra1 differentially modulate neurogenesis in neural stem cells. |journal=Autophagy |date=2012-02-01 |volume=8 |issue=2 |pages=187-99 |doi=10.4161/auto.8.2.18535 |pmid=22240590 |url=https://pubmed.ncbi.nlm.nih.gov/22240590-atg5-and-ambra1-differentially-modulate-neurogenesis-in-neural-stem-cells/ |accessdate=2020-01-28}}</ref>等過程。 |
|||
'''自噬'''({{lang-en|Autophagy}},或稱'''自體吞噬''')是一個涉及到[[細胞]]自身結構通過[[溶酶體]]機制而被[[分解代謝|分解]]的過程。這是一個受到緊密調控的步驟,此步驟是細胞生長、[[細胞分化|發育]]與[[穩態]]中的常規步驟,它幫助細胞產物在合成、降解以及接下來的循環中保持一個平衡狀態。 |
|||
自噬效應的發生取決於自噬流過程是否完成,而自噬流的意思是自噬的完整動態過程,包括自噬體形成、自噬體與溶酶體融合及後續內含物的降解和回收<ref>{{cite journal |last1=Lu |first1=Y |last2=Dong |first2=S |last3=Hao |first3=B |last4=Li |first4=C |last5=Zhu |first5=K |last6=Guo |first6=W |last7=Wang |first7=Q |last8=Cheung |first8=KH |last9=Wong |first9=CW |last10=Wu |first10=WT |last11=Markus |first11=H |last12=Yue |first12=J |title=Vacuolin-1 potently and reversibly inhibits autophagosome-lysosome fusion by activating RAB5A. |journal=Autophagy |date=2014 |volume=10 |issue=11 |pages=1895-905 |doi=10.4161/auto.32200 |pmid=25483964 |url=https://pubmed.ncbi.nlm.nih.gov/25483964-vacuolin-1-potently-and-reversibly-inhibits-autophagosome-lysosome-fusion-by-activating-rab5a/ |accessdate=2020-01-28}}</ref>。 |
|||
命名為“自噬”({{lang-en|Autophagy}})是由比利時化學家[[克里斯汀·德·迪夫]]在1963年發現的<ref name=klionsky>{{cite journal |pmid= 18567941 |year= 2008 |last1= Klionsky |first1= DJ |title= Autophagy revisited: A conversation with Christian de Duve |volume= 4 |issue= 6 |pages= 740–3 |journal= Autophagy |doi=10.4161/auto.6398}}</ref>。當代的自噬研究是1990年代酵母的研究人員通過識別的自噬相關基因而被推動<ref name="klionsky 1992">{{cite journal|pmid=1400574|title=Aminopeptidase I of Saccharomyces cerevisiae is localized to the vacuole independent of the secretory pathway|journal=Journal of Cell Biology|date=October 1992|volume=119|issue=2|page=287-99|}}</ref><ref name="ohsumi 1992">{{cite journal|pmid=1400575|title=Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction|journal=Journal of Cell Biology|date=October 1992|volume=119|issue=2|page=301-11|}}</ref><ref name="thumm 1994">{{cite journal|title=Isolation of autophagocytosis mutants of Saccharomyces cerevisiae|journal=FEBS Letters|date=August 1994|volume=349|issue=2|page=275-80|pmid=8050581}}</ref><ref name="ohsumi 1993">{{cite journal|title=Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae|journal=FEBS Letters|date=October 1993|volume=333|issue=1-2|page=169-74|pmid=8224160}}</ref><ref name="klionsky 1995">{{cite journal|title=Isolation and characterization of yeast mutants in the cytoplasm to vacuole protein targeting pathway|journal=Journal of Cell Biology|date=November 1995|volume=131|issue=3|page=591-602|pmid=7593182}}</ref>。其中之一人,日本科學家[[大隅良典]]因“對細胞自噬機制的發現”獲得2016年度的[[诺贝尔生理学或医学奖]]<ref name="nobel-2016">{{cite web|url=http://www.nobelprize.org/nobel_prizes/medicine/laureates/2016/|title=The Nobel Prize in Physiology or Medicine 2016|publisher=Nobel Foundation|accessdate=3 October 2016}}</ref>。 |
命名為“自噬”({{lang-en|Autophagy}})是由比利時化學家[[克里斯汀·德·迪夫]]在1963年發現的<ref name=klionsky>{{cite journal |pmid= 18567941 |year= 2008 |last1= Klionsky |first1= DJ |title= Autophagy revisited: A conversation with Christian de Duve |volume= 4 |issue= 6 |pages= 740–3 |journal= Autophagy |doi=10.4161/auto.6398}}</ref>。當代的自噬研究是1990年代酵母的研究人員通過識別的自噬相關基因而被推動<ref name="klionsky 1992">{{cite journal|pmid=1400574|title=Aminopeptidase I of Saccharomyces cerevisiae is localized to the vacuole independent of the secretory pathway|journal=Journal of Cell Biology|date=October 1992|volume=119|issue=2|page=287-99|}}</ref><ref name="ohsumi 1992">{{cite journal|pmid=1400575|title=Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction|journal=Journal of Cell Biology|date=October 1992|volume=119|issue=2|page=301-11|}}</ref><ref name="thumm 1994">{{cite journal|title=Isolation of autophagocytosis mutants of Saccharomyces cerevisiae|journal=FEBS Letters|date=August 1994|volume=349|issue=2|page=275-80|pmid=8050581}}</ref><ref name="ohsumi 1993">{{cite journal|title=Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae|journal=FEBS Letters|date=October 1993|volume=333|issue=1-2|page=169-74|pmid=8224160}}</ref><ref name="klionsky 1995">{{cite journal|title=Isolation and characterization of yeast mutants in the cytoplasm to vacuole protein targeting pathway|journal=Journal of Cell Biology|date=November 1995|volume=131|issue=3|page=591-602|pmid=7593182}}</ref>。其中之一人,日本科學家[[大隅良典]]因“對細胞自噬機制的發現”獲得2016年度的[[诺贝尔生理学或医学奖]]<ref name="nobel-2016">{{cite web|url=http://www.nobelprize.org/nobel_prizes/medicine/laureates/2016/|title=The Nobel Prize in Physiology or Medicine 2016|publisher=Nobel Foundation|accessdate=3 October 2016}}</ref>。 |
||
| 第120行: | 第122行: | ||
===阿茲海默症=== |
===阿茲海默症=== |
||
[[阿茲海默症]]是一種普遍流行於老年群體,並且以記憶、行為及學習功能障礙為主要特徵的神經退行性疾病<ref>{{cite journal |last1=Alzheimer’s |first1=Association. |title=2015 Alzheimer's disease facts and figures. |journal=Alzheimer's & dementia : the journal of the Alzheimer's Association |date=2015-03 |volume=11 |issue=3 |pages=332-84 |doi=10.1016/j.jalz.2015.02.003 |pmid=25984581 |url=https://pubmed.ncbi.nlm.nih.gov/25984581-2015-alzheimers-disease-facts-and-figures/ |accessdate=2020-01-25}}</ref>。自噬作用的異常是導致阿茲海默症的兩大神經病理改變,即瀰漫於整個大腦皮層的β 澱粉樣蛋白(Aβ)老年斑的形成,以及[[神經元]][[細胞核]]周圍堆積的無膜束狀異常纖維[[包涵體]](主要由過度磷酸化[[Tau蛋白]]組成)的重要原因<ref>{{cite journal |last1=Dronse |first1=J |last2=Fliessbach |first2=K |last3=Bischof |first3=GN |last4=von Reutern |first4=B |last5=Faber |first5=J |last6=Hammes |first6=J |last7=Kuhnert |first7=G |last8=Neumaier |first8=B |last9=Onur |first9=OA |last10=Kukolja |first10=J |last11=van Eimeren |first11=T |last12=Jessen |first12=F |last13=Fink |first13=GR |last14=Klockgether |first14=T |last15=Drzezga |first15=A |title=In vivo Patterns of Tau Pathology, Amyloid-β Burden, and Neuronal Dysfunction in Clinical Variants of Alzheimer's Disease. |journal=Journal of Alzheimer's disease : JAD |date=2017 |volume=55 |issue=2 |pages=465-471 |doi=10.3233/JAD-160316 |pmid=27802224 |url=https://pubmed.ncbi.nlm.nih.gov/27802224-in-vivo-patterns-of-tau-pathology-amyloid-burden-and-neuronal-dysfunction-in-clinical-variants-of-alzheimers-disease/ |accessdate=2020-01-25}}</ref>。此時的Tau蛋白失去促進微管組裝的生物學活性,並且表現出抗蛋白水解酶的神經毒性<ref>{{cite journal |last1=Ml |first1=Steinhilb |last2=D |first2=Dias-Santagata |last3=Ta |first3=Fulga |last4=Dl |first4=Felch |last5=Mb |first5=Feany |title=Tau Phosphorylation Sites Work in Concert to Promote Neurotoxicity in Vivo |journal=Molecular biology of the cell |date=2007-12 |volume=18 |issue=12 |pmid=17928404 |url=https://pubmed.ncbi.nlm.nih.gov/17928404-tau-phosphorylation-sites-work-in-concert-to-promote-neurotoxicity-in-vivo/ |accessdate=2020-01-25 |language=en}}</ref><ref>{{cite journal |last1=Mocanu |first1=MM |last2=Nissen |first2=A |last3=Eckermann |first3=K |last4=Khlistunova |first4=I |last5=Biernat |first5=J |last6=Drexler |first6=D |last7=Petrova |first7=O |last8=Schönig |first8=K |last9=Bujard |first9=H |last10=Mandelkow |first10=E |last11=Zhou |first11=L |last12=Rune |first12=G |last13=Mandelkow |first13=EM |title=The potential for beta-structure in the repeat domain of tau protein determines aggregation, synaptic decay, neuronal loss, and coassembly with endogenous Tau in inducible mouse models of tauopathy. |journal=The Journal of neuroscience : the official journal of the Society for Neuroscience |date=2008-01-16 |volume=28 |issue=3 |pages=737-48 |doi=10.1523/JNEUROSCI.2824-07.2008 |pmid=18199773 |url=https://pubmed.ncbi.nlm.nih.gov/18199773-the-potential-for-beta-structure-in-the-repeat-domain-of-tau-protein-determines-aggregation-synaptic-decay-neuronal-loss-and-coassembly-with-endogenous-tau-in-inducible-mouse-models-of-tauopathy/ |accessdate=2020-01-25}}</ref>。細胞[[囊泡]]在正常情況下,通過溶酶體途徑代謝速率很高,故而產生的Aβ量很少,不會造成Aβ的堆積 |
[[阿茲海默症]]是一種普遍流行於老年群體,並且以記憶、行為及學習功能障礙為主要特徵的神經退行性疾病<ref>{{cite journal |last1=Alzheimer’s |first1=Association. |title=2015 Alzheimer's disease facts and figures. |journal=Alzheimer's & dementia : the journal of the Alzheimer's Association |date=2015-03 |volume=11 |issue=3 |pages=332-84 |doi=10.1016/j.jalz.2015.02.003 |pmid=25984581 |url=https://pubmed.ncbi.nlm.nih.gov/25984581-2015-alzheimers-disease-facts-and-figures/ |accessdate=2020-01-25}}</ref>。自噬作用的異常是導致阿茲海默症的兩大神經病理改變,即瀰漫於整個大腦皮層的β 澱粉樣蛋白(Aβ)老年斑的形成,以及[[神經元]][[細胞核]]周圍堆積的無膜束狀異常纖維[[包涵體]](主要由過度磷酸化[[Tau蛋白]]組成)的重要原因<ref>{{cite journal |last1=Dronse |first1=J |last2=Fliessbach |first2=K |last3=Bischof |first3=GN |last4=von Reutern |first4=B |last5=Faber |first5=J |last6=Hammes |first6=J |last7=Kuhnert |first7=G |last8=Neumaier |first8=B |last9=Onur |first9=OA |last10=Kukolja |first10=J |last11=van Eimeren |first11=T |last12=Jessen |first12=F |last13=Fink |first13=GR |last14=Klockgether |first14=T |last15=Drzezga |first15=A |title=In vivo Patterns of Tau Pathology, Amyloid-β Burden, and Neuronal Dysfunction in Clinical Variants of Alzheimer's Disease. |journal=Journal of Alzheimer's disease : JAD |date=2017 |volume=55 |issue=2 |pages=465-471 |doi=10.3233/JAD-160316 |pmid=27802224 |url=https://pubmed.ncbi.nlm.nih.gov/27802224-in-vivo-patterns-of-tau-pathology-amyloid-burden-and-neuronal-dysfunction-in-clinical-variants-of-alzheimers-disease/ |accessdate=2020-01-25}}</ref>。此時的Tau蛋白失去促進微管組裝的生物學活性,並且表現出抗蛋白水解酶的神經毒性<ref>{{cite journal |last1=Ml |first1=Steinhilb |last2=D |first2=Dias-Santagata |last3=Ta |first3=Fulga |last4=Dl |first4=Felch |last5=Mb |first5=Feany |title=Tau Phosphorylation Sites Work in Concert to Promote Neurotoxicity in Vivo |journal=Molecular biology of the cell |date=2007-12 |volume=18 |issue=12 |pmid=17928404 |url=https://pubmed.ncbi.nlm.nih.gov/17928404-tau-phosphorylation-sites-work-in-concert-to-promote-neurotoxicity-in-vivo/ |accessdate=2020-01-25 |language=en}}</ref><ref>{{cite journal |last1=Mocanu |first1=MM |last2=Nissen |first2=A |last3=Eckermann |first3=K |last4=Khlistunova |first4=I |last5=Biernat |first5=J |last6=Drexler |first6=D |last7=Petrova |first7=O |last8=Schönig |first8=K |last9=Bujard |first9=H |last10=Mandelkow |first10=E |last11=Zhou |first11=L |last12=Rune |first12=G |last13=Mandelkow |first13=EM |title=The potential for beta-structure in the repeat domain of tau protein determines aggregation, synaptic decay, neuronal loss, and coassembly with endogenous Tau in inducible mouse models of tauopathy. |journal=The Journal of neuroscience : the official journal of the Society for Neuroscience |date=2008-01-16 |volume=28 |issue=3 |pages=737-48 |doi=10.1523/JNEUROSCI.2824-07.2008 |pmid=18199773 |url=https://pubmed.ncbi.nlm.nih.gov/18199773-the-potential-for-beta-structure-in-the-repeat-domain-of-tau-protein-determines-aggregation-synaptic-decay-neuronal-loss-and-coassembly-with-endogenous-tau-in-inducible-mouse-models-of-tauopathy/ |accessdate=2020-01-25}}</ref>。細胞[[囊泡]]在正常情況下,通過溶酶體途徑的代謝速率很高,故而產生的Aβ量很少,不會造成Aβ的堆積,然而自噬體轉運異常或溶酶體對自噬體內容物降解效率下降,會導致自噬體堆積、大量Aβ及老年斑的形成。Tau蛋白的兩大代謝途徑是自噬-溶酶體和泛素-蛋白體酶體系統<ref>{{cite journal |last1=Lee |first1=MJ |last2=Lee |first2=JH |last3=Rubinsztein |first3=DC |title=Tau degradation: the ubiquitin-proteasome system versus the autophagy-lysosome system. |journal=Progress in neurobiology |date=2013-06 |volume=105 |pages=49-59 |doi=10.1016/j.pneurobio.2013.03.001 |pmid=23528736 |url=https://pubmed.ncbi.nlm.nih.gov/23528736-tau-degradation-the-ubiquitin-proteasome-system-versus-the-autophagy-lysosome-system/ |accessdate=2020-01-28}}</ref>,並且是在自噬受體NDP52的協助下進行清除。自噬作用參與過度表達Tau蛋白和異常磷酸化Tau蛋白的降解,可以降低[[磷酸化]]Tau蛋白寡聚體的水平,不過對內源性Tau蛋白則没有明顯影響,並且發現抑制自噬作用能夠增加Tau蛋白的[[細胞毒性]]<ref>{{cite journal |last1=Chesser |first1=AS |last2=Ganeshan |first2=V |last3=Yang |first3=J |last4=Johnson |first4=GV |title=Epigallocatechin-3-gallate enhances clearance of phosphorylated tau in primary neurons. |journal=Nutritional neuroscience |date=2016 |volume=19 |issue=1 |pages=21-31 |doi=10.1179/1476830515Y.0000000038 |pmid=26207957 |url=https://pubmed.ncbi.nlm.nih.gov/26207957-epigallocatechin-3-gallate-enhances-clearance-of-phosphorylated-tau-in-primary-neurons/ |accessdate=2020-01-28}}</ref><ref>{{cite journal |last1=Caccamo |first1=A |last2=Magrì |first2=A |last3=Medina |first3=DX |last4=Wisely |first4=EV |last5=López-Aranda |first5=MF |last6=Silva |first6=AJ |last7=Oddo |first7=S |title=mTOR regulates tau phosphorylation and degradation: implications for Alzheimer's disease and other tauopathies. |journal=Aging cell |date=2013-06 |volume=12 |issue=3 |pages=370-80 |doi=10.1111/acel.12057 |pmid=23425014 |url=https://pubmed.ncbi.nlm.nih.gov/23425014-mtor-regulates-tau-phosphorylation-and-degradation-implications-for-alzheimers-disease-and-other-tauopathies/ |accessdate=2020-01-28}}</ref>。自噬在阿茲海默症中的作用具有兩面性。在阿茲海默症早期,由蛋白質損傷和聚集等因素誘發的自噬可以發揮保護作用。如果長[[時間]]維持這種狀態,或者溶酶體功能出現異常,就會形成[[神經元]]萎縮及細胞死亡等病理改變,所以自噬的平衡十分重要<ref>{{cite journal |last1=Cherra SJ |first1=3rd |last2=Chu |first2=CT |title=Autophagy in neuroprotection and neurodegeneration: A question of balance. |journal=Future neurology |date=2008-05 |volume=3 |issue=3 |pages=309-323 |doi=10.2217/14796708.3.3.309 |pmid=18806889 |url=https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2544613/#__ffn_sectitle |accessdate=2020-01-28}}</ref>。 |
||
===青光眼=== |
===青光眼=== |
||
2020年1月28日 (二) 05:46的版本

自噬(英語:Autophagy,或稱自體吞噬)是一個涉及到細胞自身結構通過溶酶體機制,負責將受損的細胞器、錯誤折疊的蛋白及其他大分子物質等運送至溶酶體降解並再利用的進化保守過程。自噬是廣泛存在於真核細胞的現象,並且可分為巨自噬、微自噬和分子伴侶介導的自噬三大類。這是一個受到緊密調控的步驟,此步驟是細胞生長、發育與穩態中的常規步驟,幫助細胞產物在合成、降解以及接下來的循環中保持一個平衡狀態。目前已有多份研究表明自噬在許多細胞的分化進程中被不同程度地激活[1],例如參與血管生成[2]、成骨分化[3]、脂肪生成[4]、神經發生[5]等過程。
自噬效應的發生取決於自噬流過程是否完成,而自噬流的意思是自噬的完整動態過程,包括自噬體形成、自噬體與溶酶體融合及後續內含物的降解和回收[6]。
命名為“自噬”(英語:Autophagy)是由比利時化學家克里斯汀·德·迪夫在1963年發現的[7]。當代的自噬研究是1990年代酵母的研究人員通過識別的自噬相關基因而被推動[8][9][10][11][12]。其中之一人,日本科學家大隅良典因“對細胞自噬機制的發現”獲得2016年度的诺贝尔生理学或医学奖[13]。
歷史
1962年1月,洛克菲勒醫學研究院的Keith R. Porter和他的學生Thomas Ashford報導了添加胰高血糖素後,大鼠肝細胞中的溶酶體數量增加,並且發現一些向細胞中心移位的溶酶體,包含着線粒體等細胞器的成分。Porter和Ashford錯誤地將其數據解釋為溶酶體的形成過程,不認為溶酶體是像線粒體一樣是存在於細胞質中的細胞器,並且將觀察到的水解酶理解為是由微體產生的水解酶[14]。
1963年,赫魯班(Hruban)、Spargo及其同事等報道了局部細胞質降解的超微結構,該報道參考了1955年德國科學家的損傷誘導融合模型,觀察到了從細胞質融合到生成溶酶體的三個連續步驟,並提出這個過程不僅由損傷階段誘發,而且在細胞分化的生理階段,同樣的過程也在「細胞器處置」和「細胞成分再利用」中行使功能[15]。這篇報道引起了當時也在洛克菲勒醫學研究所工作的克里斯汀·德·迪夫的興趣,與之前Porter和Ashford的看法不同,德迪夫受到這一發現的啟發,把這種現象命名為自噬(autophagy),並提出在胰高血糖素引發的肝細胞降解過程中,溶酶體發揮了功能。他與其學生拉塞爾·德特(Russell Deter)一起證實胰高血糖素誘發的自噬是由溶酶體介導的[16][17],並且在1967年連續發表兩篇文章,他也由此成為第一位報道溶酶體參與細胞內自噬的科學家。這是首次確定溶酶體是細胞內自噬的部位[7][18][19]。1974年德迪夫發現細胞內結構及功能性器官,即溶酶體和過氧物酶體,而與另外兩位科學家共享了該年度的諾貝爾生理學或醫學獎。

在1990年代,幾組科學家使用發芽酵母獨立地發現了自噬相關基因。值得注意的是,大隅良典(他於2016年獲得了諾貝爾生理學或醫學獎 ,儘管有人指出該獎項可能更具包容性[20])和Michael Thumm研究了飢餓誘導的非選擇性自噬[9][10][11]。同時,Daniel J Klionsky發現細胞質至液泡傳遞途徑(CVT),這是選擇性自噬的一種形式[8][12]。他們很快發現他們實際上是在從不同的角度看本質上相同的路徑[21][22]。最初,由酵母菌組發現的基因被賦予不同的名稱(APG、AUT、CVT、GSA、PAG、PAZ和PDD)。2003年,有研究人員提出了統一的命名法,即使用ATG表示自噬基因[23] 。
21世紀初,自噬研究領域經歷快速的發展。 ATG基因的知識為科學家提供了更方便的工具,以分析自噬在人類健康和疾病中的功能。1999年,貝絲·萊文(Beth Levine)的小組發表了一項具有里程碑意義的發現[24] ,將自噬與癌症聯繫起來。迄今為止,癌症與自噬之間的關係仍然是自噬研究的主要主題。自噬在神經退行性變和免疫防禦中的作用也受到了廣泛的關注。2003年,第一屆戈登自噬研究會議(Gordon Research Conference on autophagy)在沃特維爾舉行[25]。2005年,Daniel J Klionsky發行了致力於該領域的科學期刊《自噬》。2007年,首屆Keystone自噬專題討論會在蒙特里舉行[26]。2008年,Carol A Mercer創建了BHMT融合蛋白(GST-BHMT),該蛋白在細胞系中顯示飢餓誘導的位點特異性片段化,而甜菜鹼高半胱氨酸甲基轉移酶的降解是一種可用於評估哺乳動物細胞中自噬通量的代謝酶[27]。
巨自噬作用、微自噬作用和伴侶分子介導自噬作用由自噬相關基因及其相關酶介導[28][29][30] 。巨自噬作用細分為本體自噬和選擇性自噬(bulk and selective autophagy)。在選擇性自噬中,又細分為線粒體自噬作用[31]、脂自噬作用、過氧化物酶體自噬作用[32]、葉綠體自噬作用[33] 及核糖體自噬作用[34]等。
- 巨自噬作用是主要的自噬途徑,主要用於清除受損的細胞器或未被使用的蛋白質[35]。首先,吞噬細胞將需要降解的物質吞噬,並在受損的細胞器周圍形成自噬體[36]。然後自噬體穿過細胞的細胞質到達溶酶體,兩個細胞器融合。在溶酶體內,自噬體內的內容物通過酸性溶酶體水解酶降解[37] 。

- 伴侶分子介導自噬作用(CMA)是一個非常複雜和特異的途徑,涉及到包含hsc70的複合物的識別[36][39]。這意味著蛋白質必須包含hsc70複合物的識別位點,這將使其能夠與該分子伴侶結合,形成CMA-底物/分子伴侶複合物。然後,該複合物移動到溶酶體膜結合蛋白上,該蛋白將識別並與CMA受體結合。底物蛋白在識別後就解折疊,並在溶酶體hsc70分子伴侶的幫助下,跨越溶酶體膜轉運。CMA與其他類型的自噬存在顯著差異,因為它以一種一種的方式轉運蛋白物質,並且對哪種物質穿過溶酶體屏障具有極高的選擇性[35]。
- 線粒體自噬作用是通過自噬選擇性地降解受損或未受損的線粒體。經歷損傷或受壓後,經常發生線粒體缺陷。線粒體吞噬作用促進線粒體的更新,並且防止功能異常的線粒體積聚,從而導致細胞變性。它是由酵母中的Atg3、NIX及其調節物BNIP3在哺乳動物中介導的。線粒體吞噬作用受到PINK1和parkin蛋白的調節[40][41]。
- 脂自噬作用是通過自噬降解脂質[42],該功能在動物和真菌細胞中都存在[43]。然而,脂肪吞噬作用在植物細胞中的作用仍然難以捉摸[44]。在脂質吞噬中,靶標是稱為脂質滴(LDs)的脂質結構,具有主要是三酰基甘油(TAGs)核心,以及單層磷脂和膜蛋白組成的球形細胞器。在動物細胞中,主要的脂肪吞噬途徑是通過吞噬細胞吞噬LD,即巨自噬。另一方面,在真菌細胞中,微脂代謝是主要途徑,尤其是在發芽酵母及釀酒酵母中得到了很好的研究[45]。脂吞噬作用最早在小鼠中發現,並且在2009年發表[46]。
分子生物學
屬於絲氨酸/蘇氨酸類蛋白激酶的ATG1/ULK1是啟動自噬作用的關鍵蛋白激酶[47]。自噬的初始階段主要是誘導自噬和形成自噬膜,然而自噬膜的形成需要自噬前體(即自噬調控的重要節點)的形成。Beclin1-Vps34復合體是哺乳動物自噬的核心復合物。AtG4參與自噬泡的形成[48],而UVRAG作用於自噬泡成熟及其運輸過程[49],Rubicon負調節其功能[50]。誘導自噬後,在Atg14-Vps15-mVps34復合物作用下,啟動膜泡的成核反應,進一步結合Atg21和Atg24,形成前自噬體[51]。
自噬膜泡進一步擴張並包繞底物,最終形成自噬體。Atg12-Atg5復合物系統和LC3-Ⅱ-磷脂酰乙醇胺復合物系統均是泛素化系統,參與着自噬體的形成。目前已知p62蛋白會誘導鑲嵌有LC3的自噬體到溶酶體,將其吞噬並清除,與底物結合的p62也能被蛋白水解酶降解,而細胞內的微管骨架其實也會將自噬體運輸到溶酶體,水解酶在二者融合降解自噬體內容物,Rab7及UVRAG等因子參與此過程。目前已知Rab7與膜泡表面的脂分子尾部作用進行定位,而UVRAG則活化Rab7,將囊泡運送到靶位點[52]。
自噬過程通常涉及兩種泛素樣偶聯反應,以維持吞噬體的擴展。第1種反應發生在LC3 (酵母Atg8在哺乳動物中的同源物),LC3對溶酶體的形成是必需的。Atg4切割LC3獲得彌散胞質狀態的LC3-Ⅰ,LC3-Ⅰ其後與磷脂酰乙醇胺偶聯,以形成膜結合狀態的LC3-Ⅱ,LC3-Ⅱ其後定位於自噬體膜。Atg5-Atg12-Atg16L1復合物與前自噬體膜相關聯,通過協助招募LC3延長它們的伸長。隨著吞噬細胞擴大並接近閉合,Atg5-Atg12-Atg16L1復合物從外膜解離,而LC3-Ⅱ仍然與完成的自噬體結合。除此之外,mAtg9是核心Atg蛋白中唯一確定的多次跨膜蛋白,吞噬細胞的延伸也由mAtg9輔助。第2種反應則是在Atg12與Atg5共軛後,Atg16L1與偶聯物Atg12-Atg5結合,形成一個對吞噬細胞成熟至關重要的Atg5-Atg12-Atg16L1復合物[53]。
- ATG8酯化在自噬發生過程中的作用仍未解釋清楚,可以對促進自噬體膜的延伸和閉合起著關鍵作用。ATG8的酯化過程依賴於兩個類泛素化系統的幫助。所有生物中的ATG5-ATG12共價結合復合物對於細胞內ATG8的酯化均是必須的。
- 在自噬相關基因中,Beclin1、Atg5、Atg12和LC3對自噬的誘導階段十分重要。其中,LC3是自噬的關鍵蛋白,LC3前體正常會被ATG4切掉C端120個氨基酸,而殘片稱為LC3Ⅰ。胞膜形式的LC3Ⅰ在自噬發生時,已活化的LC3-Ⅰ經Atg3泛素化修飾後,與自噬泡膜表面的磷脂酰乙醇胺結合,形成自噬體膜形式且疏水性較強的LC3Ⅱ,並且鉚釘於自噬泡雙層膜結構上,參與細胞自噬。
- ULK復合物對自噬誘導起始至關重要[54],主要由ULK1/2、自噬相關基因13(ATG13)、ATG101和200 kD家族相互作用蛋白組成[47]。ULK1/2負責募集ATG蛋白,其中ULK1是哺乳動物自噬泡形成所必需的一種蛋白質,其活性缺失時LC3-Ⅱ不能形成,阻礙自噬過程。ULK1激酶復合體能夠促進PI3K和ATG14形成復合物,並且促進Beclin1從Bcl2-Beclin1復合體中解離出來,形成參與自噬體核形成的關鍵因子Beclin1-PI3K-ATG14復合物[48]。單磷酸腺苷活化蛋白質激酶(AMPK)和mTOR都可以催化ULK1的磷酸化[55] 。AMPK在營養充足的條件下失活,mTORC活化並磷酸化ULK1和ATG13,從而抑制自噬起始。AMPK在飢餓條件下活化,同時mTORC失活,已活化的AMPK催化ULK1絲氨酸磷酸化,促進自噬作用。除此之外,ATG13可以由mTORC1磷酸化並調控ULK復合物的活性,ATG101結合並穩定FIP200,而FIP200則為ULK及ATG13提供支架[56]。
- PI3K復合物由Vps34、Vps15、Beclin1和Atg14蛋白組成,ULK1復合物在膜泡形成後會從由細胞質轉移至內質網,並且增強PI3K復合物的活性,隨後可產生磷脂酰肌醇-3磷酸酯,促進其他效應蛋白與自噬體膜結合,從而啟動膜泡的成核反應,繼而介導前自噬體的形成[57]。
功能
營養不足
自噬在各種細胞功能中都發揮作用。營養不足會導致高水平的自噬,降解不需要的蛋白質,並且回收氨基酸,以合成對細胞生存至關重要的蛋白質[58][59][60]。在高等真核生物中,自噬作用因動物在出生後切斷了來自胎盤的食物供應而被響應[61][62]。自噬能力降低的突變酵母細胞會在營養缺乏的情況下迅速消失[63]。對APG突變體的研究表明,在飢餓條件下,通過自噬體進行的自噬對於液泡中的蛋白質降解是必不可少的,並且酵母中至少有15個APG基因參與自噬[63]。營養素介導的自噬涉及一種稱為ATG7的基因,因為小鼠研究表明,APG7缺陷型小鼠有着飢餓引起的自噬[62]。
異種吞噬
在微生物學中,異種吞噬是指感染性顆粒的自噬性降解。細胞自噬機制在先天免疫中發揮重要作用。結核桿菌等細胞內的病原體被靶向降解,當中的細胞機制和調控機制與靶向線粒體降解的機制相同[64] 。這是內共生學說的進一步證據。儘管某些細菌會阻止吞噬體成熟為降解的細胞器,稱為吞噬溶酶體,但是這個過程通常會導致由侵入性微生物帶來的破壞[65]。
感染
水皰性口炎病毒是由自噬體從胞質溶膠中吸收並轉移到內體中,並且在那裡通過TLR7檢測單鏈核糖核酸。在類鐸受體被激活後,細胞內信號級聯反應就會開始,導致干擾素和其他抗病毒細胞因子的誘導。部分病毒和細菌破壞了自噬途徑,以促進自身複製[66]。半乳凝素8已經被鑑定為細胞內的「危險受體」,能夠啟動針對細胞內病原體的自噬。當半乳凝素8與受損的液泡結合時,它會募集CALCOCO2等自噬受體,導致自噬體的形成和細菌的降解[67] 。
修復機制
自噬可以降解受損的細胞器、細胞膜和蛋白質,而抵制自噬作用被認為是造成受損細胞蓄積和衰老的其中一個主要原因[68]。自噬和自噬調節劑參與溶酶體損傷的反應,通常由半乳凝素-3和半乳凝素-8等半乳凝素指導,半乳凝素-8負責募集TRIM16。[69]和NDP52等受體,並直接影響mTOR和AMPK的活性,而mTOR和AMPK分別抑制和激活自噬作用[70]
程序性細胞死亡
程序性細胞死亡(PCD)的其中一個機制與自噬小體的出現有關,並且依賴於自噬蛋白。這種細胞死亡形式最有可能與形態學上定義為自噬PCD(autophagic PCD)的過程相對應。其中一個問題,是步入死亡過程的細胞中,其自噬的活性是導致其死亡的原因,還是為了防止細胞死亡的一個嘗試。迄今為止,形態學和組織化學研究並未證明自噬過程與細胞死亡之間存在因果關係。最近有論據認為,垂死細胞中的自噬活性可能是一種生存機制[71][72]。對昆蟲變態的研究表明,細胞經歷了一種PCD形式,這種形式與其他形式截然不同。這些已被提議作為自噬作用使細胞死亡的例子[73] 。最近的藥理和生化研究表明,有助細胞生存或致死的自噬可以通過應激期間,尤其是病毒感染後,調控信號的類型和程度來區分[74] 。然而尚未在病毒系統之外觀察到這些發現。
影響自噬的因素及其影響
力學方面
- 左心室心肌組織中,壓力超負荷會導致其蛋白酶的表達量和活性均顯著增強(目前已知蛋白酶的活性與自噬作用存在密切的關係)[75],並且導致小鼠心肌細胞的自噬活性持續地增強,甚至發現升高的情況維持至少3周[76]。許多研究證實心肌細胞的自噬活性因壓力超負荷而提高[77][78][79]。因為壓力超負荷使心肌細胞中,在細胞核周圍逐漸聚集的錯誤折疊蛋白數量增多,並且形成被細胞自噬系統識別並降解的聚集體。其後的科研人員利用動脈結扎的方法,使已經敲除Beclin-1的雜合子小鼠心臟處於壓力超負荷的狀況,最終發現敲除Beclin-1可以減弱壓力超負荷造成的細胞自噬活性提高,並且減弱心肌細胞對壓力超負荷的病理性重建作用,同時又發現在小鼠體內過度表達Beclin-1則造成相反的作用[76]。另一些科研人員進行的實驗也證明了心肌細胞為了維持細胞穩態和心肌的正常結構及功能,而擁有基礎水平的細胞自噬作用。當心肌面對着血流壓力超負荷的狀況時,自噬作用的提升就是心肌應對這種狀況的適應性反應。若然ATG5等自噬相關基因被特異性敲除,小鼠體內會出現左心室擴張及收縮功能減弱等異常現象,甚至在2周後出現了心肌肥大的問題[80]。除此之外,有多份研究指出壓力超負荷的情況消失時,細胞會逐漸回復正常的形態,而自噬則發揮著重要的作用。當去除小鼠主動脈結扎後,會發現心肌細胞逐漸復原,並且觀察到自噬相關標誌分子LC3b-Ⅱ的表達提高,反映自噬作用的活性極大地提升[81]。
- 心肌細胞若然處於去應力負荷的環境,其自噬作用會受到影響,然而自噬作用能其所促進或降低。有些研究人員心肌细胞處於去應力負荷的環境時會萎缩,並且發現LC3b-Ⅱ和Beclin-1的表達提升[82]。然而,科研人員發現有佩戴左心室輔助裝置的心臟衰竭患者,體內心肌細胞自噬的水平會降低,推測可能是因為心肌細胞的能量需求降低,不再需要維持高水平的自噬活性[83]。
- 自噬在力學刺激或衝擊引起的軟骨反應中,發揮重要的作用。最先研究有關方面的研究人員發現遭受40%力學衝擊的細胞的存活率會降低,並且發現細胞外基質中硫酸化糖胺聚糖逐漸丟失,同時在24小時後檢測到LC3b-Ⅱ的表達提高,然而在48小時後細胞的自噬逐漸受到了抑制[84]。由此可見,自噬作用在軟骨組織面對力學衝擊時,會有一定程度的保護作用,但是在損傷過大時,就會因受到抑制而失去對軟骨組織的保護作用。除此之外,有不少科研人員指出不同時程的力學刺激,對軟骨細胞的自噬影響存在着差異。例如短時程的間歇循環機械張力可以促進軟骨細胞的自噬作用,而長時程則有相反的結果[85]
- 層流剪切力對血管內皮細胞的自噬的影響在2014年已被證實。其後的研究發現層流剪切力可以促進內皮細胞的自噬活性,可能跟細胞內氧化-抗氧化平衡有關[86],而較小的層流剪切力及震蕩流剪切力卻不能夠促進自噬活性。流動剪切力(即適當的層流剪切力)能夠幫助維持內皮細胞的自噬作用,內皮細胞在加載流動剪切力後會維持靜止狀態,同時自噬的活性逐漸降低到基礎水平[87]。除此之外,有科研人員指出流動剪切力在炎症狀態下,對內皮細胞自噬的促進作用也會增強[88],並且起到抗炎症反應的作用[86]。
- 在受到機械損傷及壓縮損傷時[89],神經細胞會提升自噬的水平[90],在細胞受損早期可以透過抑制細胞凋亡的途徑來保護神經細胞。除此之外,力學因素對肝癌細胞[91]、精原瘤細胞、成纖維細胞[92]、足細胞[93]及小梁細胞等細胞自噬的促進作用也被一些研究人員所報道[94]。
鍛煉
自噬對於基礎的體內平衡至關重要,並且在鍛煉過程中保持肌肉穩態也非常重要[95][96]。對小鼠的研究表明,自噬對於不斷變化的營養和能量需求至關重要,特別是通過蛋白質進行分解代謝的代謝途徑。2012年,由德克薩斯大學達拉斯西南醫學中心進行的一項研究中,測試了具有BCL2磷酸化位點突變的小鼠驗證此理論。實驗結果顯示,這些小鼠在急性運動中,表現出耐力下降和葡萄糖代謝改變的情況[95]。另一項則研究表明,膠原蛋白VI基因敲除小鼠的骨骼肌纖維因自噬作用不足而顯示變性的跡象,導致線粒體受損和細胞死亡[97] 。運動誘導的自噬未能成功,然而在運動後以人為的方式進行自噬作用時,可以防止受損的細胞器在缺乏膠原蛋白VI的肌肉纖維中積累,並且保持細胞穩態。兩項研究均表明自噬可能有助於運動的有益代謝作用,並且在維持運動過程中的肌肉穩態,尤其是在膠原VI纖維中至關重要[95][96][97]。波恩大學細胞生物學研究所的研究表明,收縮肌肉會誘發伴侶-輔助選擇性自噬(CASA),CASA是在機械張力下維持肌肉肌節必需的[98]。CASA分子伴侶複合物識別機械損傷的細胞骨架成分,並且通過蛋白依賴性自噬途徑,將這些細胞骨架成分引導至溶酶體進行處理。這是維持肌肉活動必需的[98][99]。
炎症與自噬之間的界面
自噬調節劑控制炎症調節劑,反之亦然[100]。脊椎動物細胞通常會激活炎症,以增強免疫系統處理感染的能力,並且啟動恢復組織結構和功能的過程[101] ,故而至關重要的是將去除細胞和細菌碎片的機制與調節炎症的主要因素結合起來:溶酶體在自噬過程中對細胞成分的降解,可以回收重要的分子,並且產生及積聚物質來幫助細胞應對不斷變化的微環境[102]。控制炎症和自噬的蛋白質形成了一個對組織功能至關重要的網絡,而該網絡在癌症中失調:在癌細胞中,異常表達及突變的蛋白質增加細胞存活,對保護惡性細胞的蛋白水解系統「重新連接」網絡的依賴性[103]。這使癌細胞容易受到自噬調節劑的干預。
臨床意義

骨關節炎
由於自噬隨著年齡的增長而下降,是骨關節炎的主要危險因素,因此自噬在該疾病發展中的作用得到重視。在人類和小鼠的關節軟骨中,參與自噬的蛋白質都隨著年齡的增長而減少[104]。經培養的軟骨外植體,其機械損傷也會減少自噬蛋白[105]。自噬在正常的軟骨中會不斷被激活,但會隨著年齡的增長而受到損害,並且先於軟骨細胞死亡和出現結構受到破壞的現象[106]。因此,自噬參與了關節的正常保護過程。
炎症性腸病
炎症性腸病(inflammatory bowel disease)是由易感基因、環境和免疫系統之間一系列的相互作用所導致的慢性且易復發的消化系統疾病,包括潰瘍性結腸炎和克隆氏症等。功能失調的自噬被認為是炎症性腸病的發病因素。臨床上廣泛使用的炎症性腸病治療劑與自噬均密切相關,均能誘導細胞自噬的發生,包括類固醇、5-氨基水楊酸及硫銼嘌呤等[107]。尼古丁目前已被用作潰瘍性結腸炎患者的治療劑[108],咀嚼尼古丁口香糖可以有效控制輕度及中度結腸炎的病症[109]。經尼古丁處理後,細胞內雙層自噬泡及自噬小體的數量明顯增高。尼古丁誘導自噬的分子機制可能與內質網應激相關,或者可能與mTOR信號通路相關[110][111]。目前已有不少研究證實尼古丁與內質網應激存在密切關係,因為他們發現尼古丁上調GRP78/BIP的表達水平,並且直接誘導內質網應激,上調內質網應激標誌物PERK、EIF2A等的表達或磷酸化修飾。對低濃度尼古丁的研究有助於開發調節自噬治療潰瘍性結腸炎的新治療靶點。
牙周炎
自噬能促進感染細胞對病原體和毒素的清除,抵抗細菌的入侵。然而,牙齦卟啉單胞菌等牙周細菌則可逃避自噬分子的識別,干擾自噬體形成,阻止自噬體和溶酶體融合,甚至可以在自噬體中生存和增殖,利用其中的蛋白質等物質為自身生存提供能量[112]。目前的研究雖提示自噬與牙周炎相關,但沒有充分的證據證實自噬在牙周炎中的作用是保護作用還是病理作用。因為有研究發現牙周炎患者相較於健康人群的外周血單核細胞中,自噬基因表達量更高,而自噬作用被抑制後,出現了細胞存活率降低及凋亡細胞比例增加的情況,表明自噬在牙周炎中的保護作用[113]。然而,有研究得出相反的結果,表明自噬在牙周炎中的病理作用[114]。
敗血症及其併發症
敗血症是感染誘導的失調性宿主免疫反應。目前已知自噬在敗血症進展與器官功能障礙的發生中具有一定保護作用,故而是一個值得研究的治療靶點。大量促炎因子及聚集的白細胞通過炎症信號通路,誘導組織細胞內出現內質網應激及線粒體損傷等,繼而誘發保護性自噬反應,以包裹損傷的細胞器,限制損傷信號的擴散[115]。敗血症時,肝細胞代謝狀態會出現改變,肝細胞通過自噬來分解胞內有機物,並且產生小分子,以維持代謝需要,而分解的產物(例如糖類、脂類及氨基酸等)又可有負反饋的作用,減弱自噬作用的程度[116]。由於一氧化碳能夠增強肺部組織的自噬水平,所以有學者發表了了關於吸入低濃度一氧化碳,以治療敗血症引致的肺部損傷的報道[117]。
結核病
自噬参与到宿主细胞清除胞内致病菌的免疫应答中。結核分枝桿菌是一种胞内寄生菌,目前發現自噬有利於清除結核桿菌[118]。
在结核的免疫应答中,γ-干扰素不仅可直接加速巨噬细胞通过MHC-Ⅱ呈递抗原的作用;还可调控巨噬细胞表达IRGM,继而促进巨噬细胞内产生大量的自噬体,启动巨噬细胞的自噬作用。基因型分析发现,IRGM的某些基因型与分支杆菌的易感性明显相关,提示了IRGM基因在抗结核自噬中起到关键性作用。
除此之外,自噬也参与获得性免疫,例如加工抗原并通过MHC-Ⅱ的呈递。与此同时,一些結核桿菌的亚群也会进化出一些机制阻碍宿主细胞对其的自噬作用,达到免疫逃避的效果;有些结核杆菌可通过募集并聚合肌动蛋白以逃避泛肽化;而另一些结核杆菌则“丢卒保帅”的策略,主动脱去被泛肽化的细胞壁;这些策略都能使结核杆菌达到从吞噬体中逃离的效果。
自噬可以增強一線抗結核藥物,例如異煙肼和吡嗪酰胺的作用[119]。
癌症
當調節細胞分化的幾種不同途徑被干擾時,通常會發生癌症。自噬在癌症中起著重要作用,既可以預防癌症,也可以促進癌症的發展[120]。自噬可通過促進已飢餓或通過自噬降解凋亡介體的腫瘤細胞的存活,促進癌症的發展。自噬在癌症中的作用已得到高度研究和審查,目前已知自噬既是腫瘤抑制因子,又是腫瘤細胞存活的因素,然而根據幾種模型,自噬更可能被用作抑癌劑。在自噬的後期階段使用氯喹等抑製劑,會增加被抗腫瘤藥殺死的癌細胞數量[121]。
- 抑癌藥:目前已經對小鼠和Beclin1(一種調節自噬的蛋白質)進行了一些實驗。當Beclin1基因變為雜合子時,研究人員發現小鼠體內更容易出現腫瘤[122]。然而,當Beclin1過度表達時,腫瘤的發展就會受到抑制[123] 。在解釋beclin突變體的表型,並且將觀察結果歸因於自噬存在缺陷時應該要格外小心。Beclin1通常是產生磷脂酰肌醇3-磷酸的必需物質,因此它會影響許多溶酶體和內體功能,包括內吞作用和已活化的內吞降解生長因子受體。有認為Beclin1存在着通過非依賴自噬的途徑,影響癌症發展的可能性,然而事實是Atg7或Atg5等的核心自噬因子(暫時未知會影響其他細胞進程,並且不影響細胞增殖和細胞死亡)敲除各個基因時,顯示出非常不同的表型。此外,Beclin1的完全基因敲落會對胚胎致死,而Atg7或Atg5的敲落對則胚胎無害。壞死和慢性炎症也已顯示出通過自噬而受到限制,有助於防止腫瘤細胞的形成[124]。
- 腫瘤細胞存活率:自噬在腫瘤細胞存活中發揮重要作用。在癌細胞中,自噬被用作一種應對細胞壓力的途徑[125]。例如,miRNA-4673誘導自噬是一種有助癌細胞生存的機制,可以提高癌細胞對放射線的抵抗力[126] 。一旦這些自噬相關基因被抑制,細胞死亡的情況就會加劇[127]。自噬抵消了代謝能的增加。這些代謝壓力包括缺氧、營養缺乏及細胞增殖增加。這些壓力激活自噬,以回收ATP並維持癌細胞的存活[128]。自噬已被證明可以通過維持細胞能量的產生,而使腫瘤細胞持續地生長。通過抑制這些腫瘤細胞中的自噬基因,發現腫瘤消退,並且延長了受腫瘤影響的器官的存活率。此外,也顯示出抑制自噬可以增強抗癌治療的效果[128] 。
- 細胞死亡機制:承受極大壓力的細胞會通過細胞凋亡或壞死經歷細胞死亡。長時間的自噬激活會導致蛋白質和細胞器的高轉換率。高於生存閾值的比率,可能會殺死具有高凋亡閾值的癌細胞[128][129]。該技術可以用作癌症的治療方法。
- 治療目標:有研究發現,靶向自噬可能是抗擊癌症的可行治療方法。自噬在腫瘤抑制和腫瘤細胞存活中均起作用。因此,自噬可以用作預防癌症的策略。第一種策略是誘導自噬並增強其腫瘤抑制特性。第二種策略是抑制自噬,從而誘導細胞凋亡[127]。通過研究自噬誘導療法期間的劑量反應抗腫瘤作用,測試了第一種策略。這些療法表明自噬以劑量依賴性方式增加。 這也直接與癌細胞的生長呈劑量依賴性[125][129]。該數據支持將鼓勵自噬的療法的發展。其次,抑制直接誘導自噬的蛋白質途徑也可以用作抗癌治療[127][129]。第二種策略發現自噬是用於維持體內穩態的蛋白質降解系統,並且發現抑制自噬通常會導致細胞凋亡。抑制自噬的風險較高,因為可能導致細胞存活,而不是預計中的細胞死亡[125]。
柏金遜症

柏金遜症是一種神經退化性疾病,可以肇因於黑質緻密部的多巴胺性神經元退化。柏金遜症的特徵是在受影響的神經元中,包含着細胞無法分解的α-突觸核蛋白,其以路易氏體的形式堆積,故而柏金遜症被視為一種突觸核蛋白病變。自噬途徑的失調和調節自噬的等位基因的突變被認為會引起神經退化性疾病。自噬對神經元的生存至關重要。如果沒有有效的自噬作用,神經元會聚集遍在蛋白化的蛋白質聚集體並降解。蛋白質是已被泛素標記以降解的蛋白質。突觸核蛋白等位基因的突變導致溶酶體pH值升高和水解酶抑制。由實驗結果可知,溶酶體降解能力降低。該疾病涉及多種基因突變,包括功能喪失PTEN誘導激酶-1[131]和Parkin[132] 。這些基因的功能喪失可能導致線粒體積累和蛋白質聚集體受損,而不是導致細胞變性。線粒體參與柏金遜症。在特發性帕金森氏病中,該病通常是由線粒體功能異常、細胞氧化應激、自噬作用的改變和蛋白質聚集引起的,並會導致線粒體腫脹和去極化[133]。
阿茲海默症
阿茲海默症是一種普遍流行於老年群體,並且以記憶、行為及學習功能障礙為主要特徵的神經退行性疾病[134]。自噬作用的異常是導致阿茲海默症的兩大神經病理改變,即瀰漫於整個大腦皮層的β 澱粉樣蛋白(Aβ)老年斑的形成,以及神經元細胞核周圍堆積的無膜束狀異常纖維包涵體(主要由過度磷酸化Tau蛋白組成)的重要原因[135]。此時的Tau蛋白失去促進微管組裝的生物學活性,並且表現出抗蛋白水解酶的神經毒性[136][137]。細胞囊泡在正常情況下,通過溶酶體途徑的代謝速率很高,故而產生的Aβ量很少,不會造成Aβ的堆積,然而自噬體轉運異常或溶酶體對自噬體內容物降解效率下降,會導致自噬體堆積、大量Aβ及老年斑的形成。Tau蛋白的兩大代謝途徑是自噬-溶酶體和泛素-蛋白體酶體系統[138],並且是在自噬受體NDP52的協助下進行清除。自噬作用參與過度表達Tau蛋白和異常磷酸化Tau蛋白的降解,可以降低磷酸化Tau蛋白寡聚體的水平,不過對內源性Tau蛋白則没有明顯影響,並且發現抑制自噬作用能夠增加Tau蛋白的細胞毒性[139][140]。自噬在阿茲海默症中的作用具有兩面性。在阿茲海默症早期,由蛋白質損傷和聚集等因素誘發的自噬可以發揮保護作用。如果長時間維持這種狀態,或者溶酶體功能出現異常,就會形成神經元萎縮及細胞死亡等病理改變,所以自噬的平衡十分重要[141]。
青光眼
青光眼是一種以視網膜神經節細胞(RGCs)和視神經軸突的逐漸喪失為特徵的神經退行性疾病,並且伴有視野敏感性的喪失[142]。目前已知自噬在青光眼的發生中起着重要的作用,並且因為大量的動物實驗研究指出自噬的激活導致RGCs的凋亡,故而抑制自噬可能是預防青光眼RGCs變性的一個臨床靶點,所以有學者寄望自噬靶向治療能夠成為青光眼視神經退行性疾病的一種治療方法[143]。
病毒方面
自噬作為真核細胞早期的保護防禦機制,可以清除水疱性口炎病毒(VSV)等病原體,例如VSV的複製過程可被自噬有效抑制。然而,克沙奇病毒B3(CVB3)、丙型肝炎病毒(HCV)、乙型腦炎病毒(JEV)等病毒具有快速進化以應對宿主細胞免疫作用的能力[144],甚至會利用自噬來增強病毒的增殖[145][146][147][148]。在自噬被激活後,乙肝病毒[149]及丙肝病毒[150]會對自噬體的降解產生抑製作用。HPIV 3病毒會抑制自噬体及溶酶體的融合,阻止自噬體的降解,從而提高病毒的增殖[151]。T細胞白血病通過Tax蛋白阻斷自噬小體與溶酶體的融合,增加自噬小體的積累,並且促進病毒的複製[152][153]。有關脊髓灰質炎病毒的研究指出,當自噬體與質膜融合時,自噬體內的病毒離子被釋放[154]。
性傳播疾病方面
目前有研究指出自噬在性傳播疾病病原體的在進入宿主細胞後可以激活自噬,但是又可以逃避或抑制自噬,故而針對有關作用機制的研究或能幫助性傳播疾病的治療和預後,以及新型藥物與疫苗的研發。許多自噬相關基因都是人類免疫缺陷病毒-1感染依賴因子[155],故而HIV-1的早期複製依賴於自噬。HIV-1許多的病毒蛋白可以透過抑制自噬過程的方式,而在宿主體內持續性複製存活。例如Env編碼的外膜蛋白gp120[156]、反式激活因子Tat[157]及負性因子調控蛋白Nef[158]等。淋球菌可以抑制自噬流,增強淋球菌在巨噬細胞中的存活能力[159],也可以阻礙自噬小體成熟及自噬小體與溶酶體的融合過程,以逃避自噬介導的殺傷作用[160][161]。砂眼衣原體抑制細胞內溶酶體酶的活性,阻礙晚期自噬小體與自噬小體與溶酶體的融合過程,導致自噬的完整性受到抑制,降低自噬的抗感染作用[162]。除此之外,高危型HPV調節細胞內小分子核糖核酸的水平,影響自噬過程。例如miR-224-3p的高表達減少FIP200(參與自噬小體形成的蛋白)的水平,因抑制宿主細胞自噬而促進子宮頸癌的進展[163]。
不育症
自噬在精子發生過程中起着重要作用。精原幹細胞中的缺失或受損可引起不可逆性的弱精子症及無精子症等,導致不育症[164]。當遇精原幹細胞到不利環境時,會為了適應代謝條件的變化而進行細胞自噬調節。例如,有研究指出三鄰甲苯基磷酸酯等生殖毒性物質均可使大鼠精原幹細胞的自噬標誌蛋白及LC3-II/LC3-I 比明顯增加,細胞內含有大量退化細胞器的自噬泡顯著增加[165][166]。而且,精原幹細胞中的自噬作用可以清除錯誤折疊蛋白質,以及受損細胞器,故而對細胞起著保護作用。此外,自噬作用可以促進減數分裂前DNA複製,既為細胞提供氨基酸及核苷酸等[167][168],也保證基因遺傳的穩定性[169]。PDLIM1蛋白是一種精子細胞骨架組裝的負調控因子,並且是通過自噬途徑而降解,以維持微管結構的組裝[170]。這對精子形成有著重要作用。然而,有研究指出高脂肪食物可誘導小鼠體內的自噬作用被過度激活,導致精子生成存在缺陷[171]。
抑鬱症
自噬可能參與抑鬱症的發生[172],有多個證據提示細胞自噬的異常可能參與抑鬱症的發生[173],例如有研究指出精神分裂症斷裂基因1(一種精神疾病的關鍵易感基因)能被自噬途徑降解[174]、躁狂抑鬱症患者血清中蛋白激酶B(AKT)和mTOR 信使核糖核酸水平下降[175]、雙相抑鬱症患者腦內Bcl-2的水平下降[176],已知Bcl-2是通過與Beclin-1結合從而抑制自噬的發生。自噬對抑鬱症等神經系統疾病的調節作用是正向,抑或負向仍然存有爭議[177]。有研究提示激活自噬能促進突觸的發展[178],另一些研究則證實抗抑鬱藥可以通過抑制自噬,發揮抗抑鬱的作用。
作為藥物靶標
由於自噬失調與多種疾病的發病機理有關,因此科研人員投入了許多努力來鑑定和表徵可以調節自噬的合成人工合成小分子或天然小分子[179]。
檢測方法
第一是LC3 turnover實驗,因為單檢測LC3-II的靜態水平並不能夠完全反映細胞內的自噬潮變化,故而需要聯合自噬後期抑制劑如溶酶體抑制劑Bafilomycin A1或CQ,來比較LC3-II在抑制劑加入前後的變化差異。第二是綠色螢光蛋白(GFP)的抗降解性,通過檢測轉染了GFP-LC3的細胞所產生的GFP片段來評判細胞內自噬水平的變化。第三是以p62蛋白作為自噬活性指標,經常被科研人員用作自噬水平升高的輔助檢測手段,然而需聯合其他檢測手段進行證實。第四是mRFP-GFP-LC3雙螢光活細胞成像,實時動態監測自噬過程,並且能夠通過顏色變化確定自噬潮水平的高低。第五是使用電子顯微鏡,然而對實驗設備和實驗者的技能與辨別能力要求較高[180],有學者推薦進行雙盲實驗來定量細胞中自噬體或自噬溶酶體數量。第六是流式細胞術,可以檢測各個細胞時相的自噬水平,還可以直接計算出螢光強度和陽性細胞百分比[181],然而在細胞在染色之前,需要使用去垢劑預處理細胞質中的LC3-I。
影響因素
第一個主要影響因素是培養基的新鮮程度及血清,防止較高溫度下長時間或不適當存儲而導致的左旋麩醯胺酸(L-glutamine)降解,並產生氨。目前已知氨會因影響溶酶體的pH值而對自噬潮有明顯的抑製作用,並且通過抑制mTORC1促進自噬[182]。此外,血清也對自噬活性也有顯著影響。第二個主要影響因素是培養基的換液時間,因為培養基換液的目的,就是要減弱培養基及細胞代謝產物,對藥物靶點相關信號通路本底產生的影響。
參閱
參考文獻
- ^ Guan, JL; Simon, AK; Prescott, M; Menendez, JA; Liu, F; Wang, F; Wang, C; Wolvetang, E; Vazquez-Martin, A; Zhang, J. Autophagy in stem cells.. Autophagy. 2013-06-01, 9 (6): 830–49 [2020-01-28]. PMID 23486312. doi:10.4161/auto.24132.
- ^ Torisu, T; Torisu, K; Lee, IH; Liu, J; Malide, D; Combs, CA; Wu, XS; Rovira, II; Fergusson, MM; Weigert, R; Connelly, PS; Daniels, MP; Komatsu, M; Cao, L; Finkel, T. Autophagy regulates endothelial cell processing, maturation and secretion of von Willebrand factor.. Nature medicine. 2013-10, 19 (10): 1281–7 [2020-01-28]. PMID 24056772. doi:10.1038/nm.3288.
- ^ Pantovic, A; Krstic, A; Janjetovic, K; Kocic, J; Harhaji-Trajkovic, L; Bugarski, D; Trajkovic, V. Coordinated time-dependent modulation of AMPK/Akt/mTOR signaling and autophagy controls osteogenic differentiation of human mesenchymal stem cells.. Bone. 2013-01, 52 (1): 524–31 [2020-01-28]. PMID 23111315. doi:10.1016/j.bone.2012.10.024.
- ^ Nuschke, A; Rodrigues, M; Stolz, DB; Chu, CT; Griffith, L; Wells, A. Human mesenchymal stem cells/multipotent stromal cells consume accumulated autophagosomes early in differentiation.. Stem cell research & therapy. 2014-12-17, 5 (6): 140 [2020-01-28]. PMID 25523618. doi:10.1186/scrt530.
- ^ Vázquez, P; Arroba, AI; Cecconi, F; de la Rosa, EJ; Boya, P; de Pablo, F. Atg5 and Ambra1 differentially modulate neurogenesis in neural stem cells.. Autophagy. 2012-02-01, 8 (2): 187–99 [2020-01-28]. PMID 22240590. doi:10.4161/auto.8.2.18535.
- ^ Lu, Y; Dong, S; Hao, B; Li, C; Zhu, K; Guo, W; Wang, Q; Cheung, KH; Wong, CW; Wu, WT; Markus, H; Yue, J. Vacuolin-1 potently and reversibly inhibits autophagosome-lysosome fusion by activating RAB5A.. Autophagy. 2014, 10 (11): 1895–905 [2020-01-28]. PMID 25483964. doi:10.4161/auto.32200.
- ^ 7.0 7.1 Klionsky, DJ. Autophagy revisited: A conversation with Christian de Duve. Autophagy. 2008, 4 (6): 740–3. PMID 18567941. doi:10.4161/auto.6398.
- ^ 8.0 8.1 Aminopeptidase I of Saccharomyces cerevisiae is localized to the vacuole independent of the secretory pathway. Journal of Cell Biology. October 1992, 119 (2): 287-99. PMID 1400574.
- ^ 9.0 9.1 Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. Journal of Cell Biology. October 1992, 119 (2): 301-11. PMID 1400575.
- ^ 10.0 10.1 Isolation of autophagocytosis mutants of Saccharomyces cerevisiae. FEBS Letters. August 1994, 349 (2): 275-80. PMID 8050581.
- ^ 11.0 11.1 Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Letters. October 1993, 333 (1-2): 169-74. PMID 8224160.
- ^ 12.0 12.1 Isolation and characterization of yeast mutants in the cytoplasm to vacuole protein targeting pathway. Journal of Cell Biology. November 1995, 131 (3): 591-602. PMID 7593182.
- ^ The Nobel Prize in Physiology or Medicine 2016. Nobel Foundation. [3 October 2016].
- ^ Ashford TP, Porter KR. Cytoplasmic components in hepatic cell lysosomes. The Journal of Cell Biology. January 1962, 12 (1): 198–202. PMC 2106008
 . PMID 13862833. doi:10.1083/jcb.12.1.198.
. PMID 13862833. doi:10.1083/jcb.12.1.198.
- ^ Hruban Z, Spargo B, Swift H, Wissler RW, Kleinfeld RG. Focal cytoplasmic degradation. The American Journal of Pathology. June 1963, 42 (6): 657–83. PMC 1949709 . PMID 13955261.
- ^ Deter RL, Baudhuin P, De Duve C. Participation of lysosomes in cellular autophagy induced in rat liver by glucagon. The Journal of Cell Biology. November 1967, 35 (2): C11–6. PMC 2107130 . PMID 6055998. doi:10.1083/jcb.35.2.c11.
- ^ Deter RL, De Duve C. Influence of glucagon, an inducer of cellular autophagy, on some physical properties of rat liver lysosomes. The Journal of Cell Biology. May 1967, 33 (2): 437–49. PMC 2108350 . PMID 4292315. doi:10.1083/jcb.33.2.437.
- ^ de Duve C. Lysosomes revisited. European Journal of Biochemistry. December 1983, 137 (3): 391–7. PMID 6319122. doi:10.1111/j.1432-1033.1983.tb07841.x.
- ^ Dunn WA, Schroder LA, Aris JP. Historical overview of autophagy. Wang HG (编). Autophagy and Cancer. Springer. 2013: 3–4. ISBN 9781461465614.
- ^ Van Noorden R, Ledford H. Medicine Nobel for research on how cells 'eat themselves'. Nature. October 2016, 538 (7623): 18–19. Bibcode:2016Natur.538...18V. PMID 27708326. doi:10.1038/nature.2016.20721.
- ^ Harding TM, Hefner-Gravink A, Thumm M, Klionsky DJ. Genetic and phenotypic overlap between autophagy and the cytoplasm to vacuole protein targeting pathway. The Journal of Biological Chemistry. July 1996, 271 (30): 17621–4. PMID 8663607. doi:10.1074/jbc.271.30.17621.
- ^ Scott SV, Hefner-Gravink A, Morano KA, Noda T, Ohsumi Y, Klionsky DJ. Cytoplasm-to-vacuole targeting and autophagy employ the same machinery to deliver proteins to the yeast vacuole. Proceedings of the National Academy of Sciences of the United States of America. October 1996, 93 (22): 12304–8. Bibcode:1996PNAS...9312304S. PMC 37986 . PMID 8901576. doi:10.1073/pnas.93.22.12304.
- ^ Klionsky DJ, Cregg JM, Dunn WA, Emr SD, Sakai Y, Sandoval IV, Sibirny A, Subramani S, Thumm M, Veenhuis M, Ohsumi Y. A unified nomenclature for yeast autophagy-related genes. Developmental Cell. October 2003, 5 (4): 539–45. PMID 14536056. doi:10.1016/s1534-5807(03)00296-x.
- ^ Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, Levine B. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature. December 1999, 402 (6762): 672–6. Bibcode:1999Natur.402..672L. PMID 10604474. doi:10.1038/45257.
- ^ Autophagy in Stress, Development & Disease, 2003, Gordon Research Conference.
- ^ Autophagy in Health and Disease (Z3), 2007, Keystone Symposia on Molecular and Cellular Biology.
- ^ Mercer, CA; Kaliappan, A; Dennis, PB. Macroautophagy-dependent, intralysosomal cleavage of a betaine homocysteine methyltransferase fusion protein requires stable multimerization.. Autophagy. 2008-02, 4 (2): 185–94 [2020-01-26]. PMID 18059170. doi:10.4161/auto.5275.
- ^ Lee J, Giordano S, Zhang J. Autophagy, mitochondria and oxidative stress: cross-talk and redox signalling. The Biochemical Journal. January 2012, 441 (2): 523–40. PMC 3258656 . PMID 22187934. doi:10.1042/BJ20111451.
- ^ Mizushima N, Ohsumi Y, Yoshimori T. Autophagosome formation in mammalian cells. Cell Structure and Function. December 2002, 27 (6): 421–9. PMID 12576635. doi:10.1247/csf.27.421.
- ^ Youle RJ, Narendra DP. Mechanisms of mitophagy. Nature Reviews. Molecular Cell Biology. January 2011, 12 (1): 9–14. PMC 4780047 . PMID 21179058. doi:10.1038/nrm3028.
- ^ Ding WX, Yin XM. Mitophagy: mechanisms, pathophysiological roles, and analysis. Biological Chemistry. July 2012, 393 (7): 547–64. PMC 3630798 . PMID 22944659. doi:10.1515/hsz-2012-0119.
- ^ Till A, Lakhani R, Burnett SF, Subramani S. Pexophagy: the selective degradation of peroxisomes. International Journal of Cell Biology. 2012, 2012: 512721. PMC 3320016 . PMID 22536249. doi:10.1155/2012/512721.
- ^ Lei L. Chlorophagy: Preventing sunburn. Nature Plants. March 2017, 3 (3): 17026. PMID 28248315. doi:10.1038/nplants.2017.26.
- ^ An H, Harper JW. Systematic analysis of ribophagy in human cells reveals bystander flux during selective autophagy. Nature Cell Biology. February 2018, 20 (2): 135–143. PMC 5786475 . PMID 29230017. doi:10.1038/s41556-017-0007-x.
- ^ 35.0 35.1 Levine B, Mizushima N, Virgin HW. Autophagy in immunity and inflammation. Nature. January 2011, 469 (7330): 323–35. Bibcode:2011Natur.469..323L. PMC 3131688 . PMID 21248839. doi:10.1038/nature09782.
- ^ 36.0 36.1 36.2 Česen MH, Pegan K, Spes A, Turk B. Lysosomal pathways to cell death and their therapeutic applications. Experimental Cell Research. July 2012, 318 (11): 1245–51. PMID 22465226. doi:10.1016/j.yexcr.2012.03.005.
- ^ Homma, K.S. List of autophagy-related proteins and 3D structures. Autophagy Database. 2011, 290 [2012-10-08]. (原始内容存档于2012-08-01).
- ^ Castro-Obregon, Susana. The Discovery of Lysosomes and Autophagy. Nature Education. 2010, 3 (9): 49.
- ^ Bandyopadhyay U, Kaushik S, Varticovski L, Cuervo AM. The chaperone-mediated autophagy receptor organizes in dynamic protein complexes at the lysosomal membrane. Molecular and Cellular Biology. September 2008, 28 (18): 5747–63. PMC 2546938 . PMID 18644871. doi:10.1128/MCB.02070-07.
- ^ Gegg, ME; Cooper, JM; Chau, KY; Rojo, M; Schapira, AH; Taanman, JW. Mitofusin 1 and mitofusin 2 are ubiquitinated in a PINK1/parkin-dependent manner upon induction of mitophagy.. Human molecular genetics. 2010-12-15, 19 (24): 4861–70 [2020-01-26]. PMID 20871098. doi:10.1093/hmg/ddq419.
- ^ Geisler, S; Holmström, KM; Skujat, D; Fiesel, FC; Rothfuss, OC; Kahle, PJ; Springer, W. PINK1/Parkin-mediated mitophagy is dependent on VDAC1 and p62/SQSTM1.. Nature cell biology. 2010-02, 12 (2): 119–31 [2020-01-26]. PMID 20098416. doi:10.1038/ncb2012.
- ^ Liu K, Czaja MJ. Regulation of lipid stores and metabolism by lipophagy. Cell Death and Differentiation. January 2013, 20 (1): 3–11. PMC 3524634 . PMID 22595754. doi:10.1038/cdd.2012.63.
- ^ Ward C, Martinez-Lopez N, Otten EG, Carroll B, Maetzel D, Singh R, Sarkar S, Korolchuk VI. Autophagy, lipophagy and lysosomal lipid storage disorders. Biochimica et Biophysica Acta. April 2016, 1861 (4): 269–84. PMID 26778751. doi:10.1016/j.bbalip.2016.01.006.
- ^ Elander PH, Minina EA, Bozhkov PV. Autophagy in turnover of lipid stores: trans-kingdom comparison. Journal of Experimental Botany. March 2018, 69 (6): 1301–1311. PMID 29309625. doi:10.1093/jxb/erx433.
- ^ van Zutphen T, Todde V, de Boer R, Kreim M, Hofbauer HF, Wolinski H, Veenhuis M, van der Klei IJ, Kohlwein SD. Lipid droplet autophagy in the yeast Saccharomyces cerevisiae. Molecular Biology of the Cell. January 2014, 25 (2): 290–301. PMC 3890349 . PMID 24258026. doi:10.1091/mbc.E13-08-0448.
- ^ Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, Komatsu M, Tanaka K, Cuervo AM, Czaja MJ. Autophagy regulates lipid metabolism. Nature. April 2009, 458 (7242): 1131–5. Bibcode:2009Natur.458.1131S. PMC 2676208 . PMID 19339967. doi:10.1038/nature07976.
- ^ 47.0 47.1 Hosokawa, N; Hara, T; Kaizuka, T; Kishi, C; Takamura, A; Miura, Y; Iemura, S; Natsume, T; Takehana, K; Yamada, N; Guan, JL; Oshiro, N; Mizushima, N. Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy.. Molecular biology of the cell. 2009-04, 20 (7): 1981–91 [2020-01-26]. PMID 19211835. doi:10.1091/mbc.e08-12-1248.
- ^ 48.0 48.1 Itakura, E; Kishi, C; Inoue, K; Mizushima, N. Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG.. Molecular biology of the cell. 2008-12, 19 (12): 5360–72 [2020-01-26]. PMID 18843052. doi:10.1091/mbc.e08-01-0080.
- ^ Liang, C; Lee, JS; Inn, KS; Gack, MU; Li, Q; Roberts, EA; Vergne, I; Deretic, V; Feng, P; Akazawa, C; Jung, JU. Beclin1-binding UVRAG targets the class C Vps complex to coordinate autophagosome maturation and endocytic trafficking.. Nature cell biology. 2008-07, 10 (7): 776–87 [2020-01-26]. PMID 18552835. doi:10.1038/ncb1740.
- ^ Matsunaga, K; Saitoh, T; Tabata, K; Omori, H; Satoh, T; Kurotori, N; Maejima, I; Shirahama-Noda, K; Ichimura, T; Isobe, T; Akira, S; Noda, T; Yoshimori, T. Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages.. Nature cell biology. 2009-04, 11 (4): 385–96 [2020-01-26]. PMID 19270696. doi:10.1038/ncb1846.
- ^ He, C; Klionsky, DJ. Regulation mechanisms and signaling pathways of autophagy.. Annual review of genetics. 2009, 43: 67–93 [2020-01-26]. PMID 19653858. doi:10.1146/annurev-genet-102808-114910.
- ^ Luo, P; Gao, F; Han, J; Sun, W; Li, Z. The role of autophagy in steroid necrosis of the femoral head: a comprehensive research review.. International orthopaedics. 2018-07, 42 (7): 1747–1753 [2020-01-26]. PMID 29797168. doi:10.1007/s00264-018-3994-8.
- ^ Rubinsztein, DC; Shpilka, T; Elazar, Z. Mechanisms of autophagosome biogenesis.. Current biology : CB. 2012-01-10, 22 (1): R29–34 [2020-01-26]. PMID 22240478. doi:10.1016/j.cub.2011.11.034.
- ^ Matsuura, A; Tsukada, M; Wada, Y; Ohsumi, Y. Apg1p, a novel protein kinase required for the autophagic process in Saccharomyces cerevisiae.. Gene. 1997-06-19, 192 (2): 245–50 [2020-01-26]. PMID 9224897. doi:10.1016/s0378-1119(97)00084-x.
- ^ Chan EY. Regulation and function of uncoordinated-51 like kinase proteins. Antioxidants & Redox Signaling. September 2012, 17 (5): 775–85. PMID 22074133. doi:10.1089/ars.2011.4396.
- ^ Choi, Y; Bowman, JW; Jung, JU. Autophagy during viral infection - a double-edged sword.. Nature reviews. Microbiology. 2018-06, 16 (6): 341–354 [2020-01-26]. PMID 29556036. doi:10.1038/s41579-018-0003-6.
- ^ Ohsumi, Y. Historical landmarks of autophagy research.. Cell research. 2014-01, 24 (1): 9–23 [2020-01-26]. PMID 24366340. doi:10.1038/cr.2013.169.
- ^ Reggiori F, Klionsky DJ. Autophagy in the eukaryotic cell. Eukaryotic Cell. February 2002, 1 (1): 11–21. PMC 118053 . PMID 12455967. doi:10.1128/EC.01.1.11-21.2002.
- ^ Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation. Science. December 2000, 290 (5497): 1717–21. Bibcode:2000Sci...290.1717K. PMC 2732363 . PMID 11099404. doi:10.1126/science.290.5497.1717.
- ^ Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Developmental Cell. April 2004, 6 (4): 463–77. PMID 15068787. doi:10.1016/S1534-5807(04)00099-1.
- ^ Kuma A, Hatano M, Matsui M, Yamamoto A, Nakaya H, Yoshimori T, Ohsumi Y, Tokuhisa T, Mizushima N, et al. The role of autophagy during the early neonatal starvation period. Nature. December 2004, 432 (7020): 1032–6. Bibcode:2004Natur.432.1032K. PMID 15525940. doi:10.1038/nature03029.
- ^ 62.0 62.1 Mizushima N, Yamamoto A, Matsui M, Yoshimori T, Ohsumi Y. In vivo analysis of autophagy in response to nutrient starvation using transgenic mice expressing a fluorescent autophagosome marker. Molecular Biology of the Cell. March 2004, 15 (3): 1101–11. PMC 363084 . PMID 14699058. doi:10.1091/mbc.E03-09-0704.
- ^ 63.0 63.1 Tsukada M, Ohsumi Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Letters. October 1993, 333 (1–2): 169–74. PMID 8224160. doi:10.1016/0014-5793(93)80398-E.
- ^ Gutierrez MG, Master SS, Singh SB, Taylor GA, Colombo MI, Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell. December 2004, 119 (6): 753–66. PMID 15607973. doi:10.1016/j.cell.2004.11.038.
- ^ Deretic V, Delgado M, Vergne I, Master S, De Haro S, Ponpuak M, Singh S. Autophagy in immunity against mycobacterium tuberculosis: a model system to dissect immunological roles of autophagy. Autophagy in Infection and Immunity. Current Topics in Microbiology and Immunology 335. 2009: 169–88. ISBN 978-3-642-00301-1. PMC 2788935 . PMID 19802565. doi:10.1007/978-3-642-00302-8_8.
- ^ Jackson WT, Giddings TH, Taylor MP, Mulinyawe S, Rabinovitch M, Kopito RR, Kirkegaard K. Subversion of cellular autophagosomal machinery by RNA viruses. PLoS Biology. May 2005, 3 (5): e156. PMC 1084330 . PMID 15884975. doi:10.1371/journal.pbio.0030156.

- ^ Thurston TL, Wandel MP, von Muhlinen N, Foeglein A, Randow F. Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature. January 2012, 482 (7385): 414–8. Bibcode:2012Natur.482..414T. PMC 3343631 . PMID 22246324. doi:10.1038/nature10744.
- ^ Cuervo AM, Bergamini E, Brunk UT, Dröge W, Ffrench M, Terman A. Autophagy and aging: the importance of maintaining "clean" cells. Autophagy. 2005, 1 (3): 131–40. PMID 16874025. doi:10.4161/auto.1.3.2017.
- ^ Chauhan S, Kumar S, Jain A, Ponpuak M, Mudd MH, Kimura T, Choi SW, Peters R, Mandell M, Bruun JA, Johansen T, Deretic V. TRIMs and Galectins Globally Cooperate and TRIM16 and Galectin-3 Co-direct Autophagy in Endomembrane Damage Homeostasis. Developmental Cell. October 2016, 39 (1): 13–27. PMC 5104201 . PMID 27693506. doi:10.1016/j.devcel.2016.08.003.
- ^ Jia J, Abudu YP, Claude-Taupin A, Gu Y, Kumar S, Choi SW, Peters R, Mudd MH, Allers L, Salemi M, Phinney B, Johansen T, Deretic V. Galectins Control mTOR in Response to Endomembrane Damage. Molecular Cell. April 2018, 70 (1): 120–135.e8. PMC 5911935 . PMID 29625033. doi:10.1016/j.molcel.2018.03.009 (英语).
- ^ Tavassoly I. Dynamics of Cell Fate Decision Mediated by the Interplay of Autophagy and Apoptosis in Cancer Cells. Springer Theses. Springer International Publishing. 2015. ISBN 978-3-319-14962-2. doi:10.1007/978-3-319-14962-2.
- ^ Tsujimoto Y, Shimizu S. Another way to die: autophagic programmed cell death. Cell Death and Differentiation. November 2005,. 12 Suppl 2 (Suppl 2): 1528–34. PMID 16247500. doi:10.1038/sj.cdd.4401777.
- ^ Schwartz LM, Smith SW, Jones ME, Osborne BA. Do all programmed cell deaths occur via apoptosis?. Proceedings of the National Academy of Sciences of the United States of America. February 1993, 90 (3): 980–4. Bibcode:1993PNAS...90..980S. PMC 45794 . PMID 8430112. doi:10.1073/pnas.90.3.980.
- ^ Datan E, Shirazian A, Benjamin S, Matassov D, Tinari A, Malorni W, Lockshin RA, Garcia-Sastre A, Zakeri Z. mTOR/p70S6K signaling distinguishes routine, maintenance-level autophagy from autophagic cell death during influenza A infection. Virology. March 2014,. 452-453 (March 2014): 175–190. PMC 4005847 . PMID 24606695. doi:10.1016/j.virol.2014.01.008.
- ^ Depre, C; Wang, Q; Yan, L; Hedhli, N; Peter, P; Chen, L; Hong, C; Hittinger, L; Ghaleh, B; Sadoshima, J; Vatner, DE; Vatner, SF; Madura, K. Activation of the cardiac proteasome during pressure overload promotes ventricular hypertrophy.. Circulation. 2006-10-24, 114 (17): 1821–8 [2020-01-25]. PMID 17043166. doi:10.1161/CIRCULATIONAHA.106.637827.
- ^ 76.0 76.1 Zhu, H; Tannous, P; Johnstone, JL; Kong, Y; Shelton, JM; Richardson, JA; Le, V; Levine, B; Rothermel, BA; Hill, JA. Cardiac autophagy is a maladaptive response to hemodynamic stress.. The Journal of clinical investigation. 2007-07, 117 (7): 1782–93 [2020-01-25]. PMID 17607355. doi:10.1172/JCI27523.
- ^ Fu, L; Wei, CC; Powell, PC; Bradley, WE; Collawn, JF; Dell'Italia, LJ. Volume overload induces autophagic degradation of procollagen in cardiac fibroblasts.. Journal of molecular and cellular cardiology. 2015-12, 89 (Pt B): 241–250 [2020-01-25]. PMID 26596413. doi:10.1016/j.yjmcc.2015.10.027.
- ^ Lin, L; Liu, X; Xu, J; Weng, L; Ren, J; Ge, J; Zou, Y. High-density lipoprotein inhibits mechanical stress-induced cardiomyocyte autophagy and cardiac hypertrophy through angiotensin II type 1 receptor-mediated PI3K/Akt pathway.. Journal of cellular and molecular medicine. 2015-08, 19 (8): 1929–38 [2020-01-25]. PMID 25946687. doi:10.1111/jcmm.12567.
- ^ Weng, LQ; Zhang, WB; Ye, Y; Yin, PP; Yuan, J; Wang, XX; Kang, L; Jiang, SS; You, JY; Wu, J; Gong, H; Ge, JB; Zou, YZ. Aliskiren ameliorates pressure overload-induced heart hypertrophy and fibrosis in mice.. Acta pharmacologica Sinica. 2014-08, 35 (8): 1005–14 [2020-01-25]. PMID 24998254. doi:10.1038/aps.2014.45.
- ^ Nakai, A; Yamaguchi, O; Takeda, T; Higuchi, Y; Hikoso, S; Taniike, M; Omiya, S; Mizote, I; Matsumura, Y; Asahi, M; Nishida, K; Hori, M; Mizushima, N; Otsu, K. The role of autophagy in cardiomyocytes in the basal state and in response to hemodynamic stress.. Nature medicine. 2007-05, 13 (5): 619–24 [2020-01-25]. PMID 17450150. doi:10.1038/nm1574.
- ^ Hariharan, N; Ikeda, Y; Hong, C; Alcendor, RR; Usui, S; Gao, S; Maejima, Y; Sadoshima, J. Autophagy plays an essential role in mediating regression of hypertrophy during unloading of the heart.. PloS one. 2013, 8 (1): e51632 [2020-01-25]. PMID 23308102. doi:10.1371/journal.pone.0051632.
- ^ Cao, DJ; Jiang, N; Blagg, A; Johnstone, JL; Gondalia, R; Oh, M; Luo, X; Yang, KC; Shelton, JM; Rothermel, BA; Gillette, TG; Dorn, GW; Hill, JA. Mechanical unloading activates FoxO3 to trigger Bnip3-dependent cardiomyocyte atrophy.. Journal of the American Heart Association. 2013-04-08, 2 (2): e000016 [2020-01-25]. PMID 23568341. doi:10.1161/JAHA.113.000016.
- ^ Kassiotis, C; Ballal, K; Wellnitz, K; Vela, D; Gong, M; Salazar, R; Frazier, OH; Taegtmeyer, H. Markers of autophagy are downregulated in failing human heart after mechanical unloading.. Circulation. 2009-09-15, 120 (11 Suppl): S191–7 [2020-01-25]. PMID 19752367. doi:10.1161/CIRCULATIONAHA.108.842252.
- ^ Caramés, B; Taniguchi, N; Seino, D; Blanco, FJ; D'Lima, D; Lotz, M. Mechanical injury suppresses autophagy regulators and pharmacologic activation of autophagy results in chondroprotection.. Arthritis and rheumatism. 2012-04, 64 (4): 1182–92 [2020-01-25]. PMID 22034068. doi:10.1002/art.33444.
- ^ Xu, Hong-guang; Yu, Yun-fei; Zheng, Quan; Zhang, Wei; Wang, Chuang-dong; Zhao, Xiao-yin; Tong, Wen-xue; Wang, Hong; Liu, Ping; Zhang, Xiao-ling. Autophagy protects end plate chondrocytes from intermittent cyclic mechanical tension induced calcification. Bone. 2014-09, 66: 232–239. doi:10.1016/j.bone.2014.06.018.
- ^ 86.0 86.1 Bharath, LP; Mueller, R; Li, Y; Ruan, T; Kunz, D; Goodrich, R; Mills, T; Deeter, L; Sargsyan, A; Anandh Babu, PV; Graham, TE; Symons, JD. Impairment of autophagy in endothelial cells prevents shear-stress-induced increases in nitric oxide bioavailability.. Canadian journal of physiology and pharmacology. 2014-07, 92 (7): 605–12 [2020-01-25]. PMID 24941409. doi:10.1139/cjpp-2014-0017.
- ^ Guo, F; Li, X; Peng, J; Tang, Y; Yang, Q; Liu, L; Wang, Z; Jiang, Z; Xiao, M; Ni, C; Chen, R; Wei, D; Wang, GX. Autophagy regulates vascular endothelial cell eNOS and ET-1 expression induced by laminar shear stress in an ex vivo perfused system.. Annals of biomedical engineering. 2014-09, 42 (9): 1978–88 [2020-01-25]. PMID 24838486. doi:10.1007/s10439-014-1033-5.
- ^ Ding, Z; Liu, S; Deng, X; Fan, Y; Wang, X; Mehta, JL. Hemodynamic shear stress modulates endothelial cell autophagy: Role of LOX-1.. International journal of cardiology. 2015-04-01, 184: 86–95 [2020-01-25]. PMID 25697875. doi:10.1016/j.ijcard.2015.01.065.
- ^ Chen, Z; Fu, Q; Shen, B; Huang, X; Wang, K; He, P; Li, F; Zhang, F; Shen, H. Enhanced p62 expression triggers concomitant autophagy and apoptosis in a rat chronic spinal cord compression model.. Molecular medicine reports. 2014-06, 9 (6): 2091–6 [2020-01-25]. PMID 24715058. doi:10.3892/mmr.2014.2124.
- ^ Chen, HC; Fong, TH; Hsu, PW; Chiu, WT. Multifaceted effects of rapamycin on functional recovery after spinal cord injury in rats through autophagy promotion, anti-inflammation, and neuroprotection.. The Journal of surgical research. 2013-01, 179 (1): e203–10 [2020-01-25]. PMID 22482761. doi:10.1016/j.jss.2012.02.023.
- ^ Lien, SC; Chang, SF; Lee, PL; Wei, SY; Chang, MD; Chang, JY; Chiu, JJ. Mechanical regulation of cancer cell apoptosis and autophagy: roles of bone morphogenetic protein receptor, Smad1/5, and p38 MAPK.. Biochimica et biophysica acta. 2013-12, 1833 (12): 3124–3133 [2020-01-25]. PMID 24021264. doi:10.1016/j.bbamcr.2013.08.023.
- ^ Chen, H; Chen, L; Cheng, B; Jiang, C. Cyclic mechanical stretching induces autophagic cell death in tenofibroblasts through activation of prostaglandin E2 production.. Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology. 2015, 36 (1): 24–33 [2020-01-25]. PMID 25924624. doi:10.1159/000374050.
- ^ Li, D; Lu, Z; Xu, Z; Ji, J; Zheng, Z; Lin, S; Yan, T. Spironolactone promotes autophagy via inhibiting PI3K/AKT/mTOR signalling pathway and reduce adhesive capacity damage in podocytes under mechanical stress.. Bioscience reports. 2016-08, 36 (4) [2020-01-25]. PMID 27129295. doi:10.1042/BSR20160086.
- ^ Porter, KM; Jeyabalan, N; Liton, PB. MTOR-independent induction of autophagy in trabecular meshwork cells subjected to biaxial stretch.. Biochimica et biophysica acta. 2014-06, 1843 (6): 1054–62 [2020-01-25]. PMID 24583119. doi:10.1016/j.bbamcr.2014.02.010.
- ^ 95.0 95.1 95.2 He C, Bassik MC, Moresi V, Sun K, Wei Y, Zou Z, An Z, Loh J, Fisher J, Sun Q, Korsmeyer S, Packer M, May HI, Hill JA, Virgin HW, Gilpin C, Xiao G, Bassel-Duby R, Scherer PE, Levine B, et al. Exercise-induced BCL2-regulated autophagy is required for muscle glucose homeostasis. Nature. January 2012, 481 (7382): 511–5. Bibcode:2012Natur.481..511H. PMC 3518436 . PMID 22258505. doi:10.1038/nature10758.
- ^ 96.0 96.1 Nair U, Klionsky DJ. Activation of autophagy is required for muscle homeostasis during physical exercise. Autophagy. December 2011, 7 (12): 1405–6. PMC 3288013 . PMID 22082869. doi:10.4161/auto.7.12.18315.
- ^ 97.0 97.1 Grumati P, Coletto L, Schiavinato A, Castagnaro S, Bertaggia E, Sandri M, Bonaldo P. Physical exercise stimulates autophagy in normal skeletal muscles but is detrimental for collagen VI-deficient muscles. Autophagy. December 2011, 7 (12): 1415–23. PMC 3288016 . PMID 22024752. doi:10.4161/auto.7.12.17877.
- ^ 98.0 98.1 Arndt V, Dick N, Tawo R, Dreiseidler M, Wenzel D, Hesse M, Fürst DO, Saftig P, Saint R, Fleischmann BK, Hoch M, Höhfeld J. Chaperone-assisted selective autophagy is essential for muscle maintenance. Current Biology. January 2010, 20 (2): 143–8. PMID 20060297. doi:10.1016/j.cub.2009.11.022.
- ^ Ulbricht A, Eppler FJ, Tapia VE, van der Ven PF, Hampe N, Hersch N, Vakeel P, Stadel D, Haas A, Saftig P, Behrends C, Fürst DO, Volkmer R, Hoffmann B, Kolanus W, Höhfeld J. Cellular mechanotransduction relies on tension-induced and chaperone-assisted autophagy. Current Biology. March 2013, 23 (5): 430–5. PMID 23434281. doi:10.1016/j.cub.2013.01.064.
- ^ Cadwell, K. Crosstalk between autophagy and inflammatory signalling pathways: balancing defence and homeostasis.. Nature. 2016, 16 (11): 661–675. PMC 5343289 . PMID 27694913. doi:10.1038/nri.2016.100.
- ^ Medzhitov, R. Origin and physiological roles of inflammation.. Nature. 2008, 454 (7203): 428–435. PMID 18650913. doi:10.1038/nature07201.
- ^ Tan, P. Autophagy and Immune-Related Diseases.. Adv Exp Med Biol. Advances in Experimental Medicine and Biology. 2019, 1209: 167–179. ISBN 978-981-15-0605-5. PMID 31728870. doi:10.1007/978-981-15-0606-2_10.
- ^ Varisli, L. Dissecting pharmacological effects of Chloroquine in cancer treatment: interference with inflammatory signaling pathways.. Immunology. 2019. PMID 31782148. doi:10.1111/imm.13160.
- ^ Caramés B, Taniguchi N, Otsuki S, Blanco FJ, Lotz M. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis and Rheumatism. March 2010, 62 (3): 791–801. PMC 2838960 . PMID 20187128. doi:10.1002/art.27305.
- ^ Caramés B, Taniguchi N, Seino D, Blanco FJ, D'Lima D, Lotz M. Mechanical injury suppresses autophagy regulators and pharmacologic activation of autophagy results in chondroprotection. Arthritis and Rheumatism. April 2012, 64 (4): 1182–92. PMC 3288456 . PMID 22034068. doi:10.1002/art.33444.
- ^ Caramés B, Olmer M, Kiosses WB, Lotz MK. The relationship of autophagy defects to cartilage damage during joint aging in a mouse model. Arthritis & Rheumatology. June 2015, 67 (6): 1568–76. PMC 4446178 . PMID 25708836. doi:10.1002/art.39073.
- ^ Iida, T; Onodera, K; Nakase, H. Role of autophagy in the pathogenesis of inflammatory bowel disease.. World journal of gastroenterology. 2017-03-21, 23 (11): 1944–1953 [2020-01-23]. PMID 28373760. doi:10.3748/wjg.v23.i11.1944.
- ^ Lunney, PC; Leong, RW. Review article: Ulcerative colitis, smoking and nicotine therapy.. Alimentary pharmacology & therapeutics. 2012-12, 36 (11-12): 997–1008 [2020-01-23]. PMID 23072629. doi:10.1111/apt.12086.
- ^ Berkowitz, L; Schultz, BM; Salazar, GA; Pardo-Roa, C; Sebastián, VP; Álvarez-Lobos, MM; Bueno, SM. Impact of Cigarette Smoking on the Gastrointestinal Tract Inflammation: Opposing Effects in Crohn's Disease and Ulcerative Colitis.. Frontiers in immunology. 2018, 9: 74 [2020-01-23]. PMID 29441064. doi:10.3389/fimmu.2018.00074.
- ^ Wong, MK; Holloway, AC; Hardy, DB. Nicotine Directly Induces Endoplasmic Reticulum Stress Response in Rat Placental Trophoblast Giant Cells.. Toxicological sciences : an official journal of the Society of Toxicology. 2016-05, 151 (1): 23–34 [2020-01-23]. PMID 26803847. doi:10.1093/toxsci/kfw019.
- ^ Guan, Y; Zhang, L; Li, X; Zhang, X; Liu, S; Gao, N; Li, L; Gao, G; Wei, G; Chen, Z; Zheng, Y; Ma, X; Siwko, S; Chen, JL; Liu, M; Li, D. Repression of Mammalian Target of Rapamycin Complex 1 Inhibits Intestinal Regeneration in Acute Inflammatory Bowel Disease Models.. Journal of immunology (Baltimore, Md. : 1950). 2015-07-01, 195 (1): 339–46 [2020-01-23]. PMID 26026060. doi:10.4049/jimmunol.1303356.
- ^ Bélanger, M; Rodrigues, PH; Dunn WA, Jr; Progulske-Fox, A. Autophagy: a highway for Porphyromonas gingivalis in endothelial cells.. Autophagy. NaN, 2 (3): 165–70 [2020-01-23]. PMID 16874051. doi:10.4161/auto.2828.
- ^ Levine, B; Klionsky, DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy.. Developmental cell. 2004-04, 6 (4): 463–77 [2020-01-23]. PMID 15068787. doi:10.1016/s1534-5807(04)00099-1.
- ^ Tsuda, H; Ochiai, K; Suzuki, N; Otsuka, K. Butyrate, a bacterial metabolite, induces apoptosis and autophagic cell death in gingival epithelial cells.. Journal of periodontal research. 2010-10, 45 (5): 626–34 [2020-01-23]. PMID 20546110. doi:10.1111/j.1600-0765.2010.01277.x.
- ^ Lewis, AJ; Billiar, TR; Rosengart, MR. Biology and Metabolism of Sepsis: Innate Immunity, Bioenergetics, and Autophagy.. Surgical infections. 2016-06, 17 (3): 286–93 [2020-01-25]. PMID 27093228. doi:10.1089/sur.2015.262.
- ^ Madrigal-Matute, J; Cuervo, AM. Regulation of Liver Metabolism by Autophagy.. Gastroenterology. 2016-02, 150 (2): 328–39 [2020-01-25]. PMID 26453774. doi:10.1053/j.gastro.2015.09.042.
- ^ Hoetzel, A; Dolinay, T; Schmidt, R; Choi, AM; Ryter, SW. Carbon monoxide in sepsis.. Antioxidants & redox signaling. 2007-11, 9 (11): 2013–26 [2020-01-25]. PMID 17822362. doi:10.1089/ars.2007.1762.
- ^ Chen, S; Yuan, J; Yao, S; Jin, Y; Chen, G; Tian, W; Xi, J; Xu, Z; Weng, D; Chen, J. Lipopolysaccharides may aggravate apoptosis through accumulation of autophagosomes in alveolar macrophages of human silicosis.. Autophagy. 2015, 11 (12): 2346–57 [2020-01-23]. PMID 26553601. doi:10.1080/15548627.2015.1109765.
- ^ Kim, JJ; Lee, HM; Shin, DM; Kim, W; Yuk, JM; Jin, HS; Lee, SH; Cha, GH; Kim, JM; Lee, ZW; Shin, SJ; Yoo, H; Park, YK; Park, JB; Chung, J; Yoshimori, T; Jo, EK. Host cell autophagy activated by antibiotics is required for their effective antimycobacterial drug action.. Cell host & microbe. 2012-05-17, 11 (5): 457–68 [2020-01-23]. PMID 22607799. doi:10.1016/j.chom.2012.03.008.
- ^ Furuya, N., Liang, X.H., and Levin, B. 2004. Autophagy and cancer. In Autophagy. D.J. Klionsky editor. Landes Bioscience. Georgetown, Texas, USA. 244-253.
- ^ Vlahopoulos S, Critselis E, Voutsas IF, Perez SA, Moschovi M, Baxevanis CN, Chrousos GP. New use for old drugs? Prospective targets of chloroquines in cancer therapy. Current Drug Targets. 2014, 15 (9): 843–51. PMID 25023646. doi:10.2174/1389450115666140714121514.
- ^ Qu X, Yu J, Bhagat G, Furuya N, Hibshoosh H, Troxel A, Rosen J, Eskelinen EL, Mizushima N, Ohsumi Y, Cattoretti G, Levine B, et al. Promotion of tumorigenesis by heterozygous disruption of the beclin 1 autophagy gene. The Journal of Clinical Investigation. December 2003, 112 (12): 1809–20. PMC 297002 . PMID 14638851. doi:10.1172/JCI20039.
- ^ Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, Levine B, et al. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature. December 1999, 402 (6762): 672–6. Bibcode:1999Natur.402..672L. PMID 10604474. doi:10.1038/45257.
- ^ Duran A, Linares JF, Galvez AS, Wikenheiser K, Flores JM, Diaz-Meco MT, Moscat J, et al. The signaling adaptor p62 is an important NF-kappaB mediator in tumorigenesis. Cancer Cell. April 2008, 13 (4): 343–54. PMID 18394557. doi:10.1016/j.ccr.2008.02.001.
- ^ 125.0 125.1 125.2 Paglin S, Hollister T, Delohery T, Hackett N, McMahill M, Sphicas E, Domingo D, Yahalom J. A novel response of cancer cells to radiation involves autophagy and formation of acidic vesicles. Cancer Research. January 2001, 61 (2): 439–44. PMID 11212227.
- ^ Dökümcü K, Simonian M, Farahani RM. miR4673 improves fitness profile of neoplastic cells by induction of autophagy. Cell Death & Disease. October 2018, 9 (11): 1068. PMC 6195512 . PMID 30341280. doi:10.1038/s41419-018-1088-6.
- ^ 127.0 127.1 127.2 Jin S, White E. Role of autophagy in cancer: management of metabolic stress. Autophagy. 2007, 3 (1): 28–31. PMC 2770734 . PMID 16969128. doi:10.4161/auto.3269.
- ^ 128.0 128.1 128.2 Yang ZJ, Chee CE, Huang S, Sinicrope FA. The role of autophagy in cancer: therapeutic implications. Molecular Cancer Therapeutics. September 2011, 10 (9): 1533–41. PMC 3170456 . PMID 21878654. doi:10.1158/1535-7163.MCT-11-0047.
- ^ 129.0 129.1 129.2 Tavassoly I, Parmar J, Shajahan-Haq AN, Clarke R, Baumann WT, Tyson JJ. Dynamic Modeling of the Interaction Between Autophagy and Apoptosis in Mammalian Cells. CPT. April 2015, 4 (4): 263–72. PMC 4429580 . PMID 26225250. doi:10.1002/psp4.29.
- ^ Razaghi A, Heimann K, Schaeffer PM, Gibson SB. Negative regulators of cell death pathways in cancer: perspective on biomarkers and targeted therapies. Apoptosis. February 2018, 23 (2): 93–112. PMID 29322476. doi:10.1007/s10495-018-1440-4.
- ^ Valente EM, Abou-Sleiman PM, Caputo V, Muqit MM, Harvey K, Gispert S, Ali Z, Del Turco D, Bentivoglio AR, Healy DG, Albanese A, Nussbaum R, González-Maldonado R, Deller T, Salvi S, Cortelli P, Gilks WP, Latchman DS, Harvey RJ, Dallapiccola B, Auburger G, Wood NW. Hereditary early-onset Parkinson's disease caused by mutations in PINK1. Science. May 2004, 304 (5674): 1158–60. Bibcode:2004Sci...304.1158V. PMID 15087508. doi:10.1126/science.1096284.
- ^ Kitada T, Asakawa S, Hattori N, Matsumine H, Yamamura Y, Minoshima S, Yokochi M, Mizuno Y, Shimizu N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature. April 1998, 392 (6676): 605–8. Bibcode:1998Natur.392..605K. PMID 9560156. doi:10.1038/33416.
- ^ Esteves AR, Arduíno DM, Silva DF, Oliveira CR, Cardoso SM. Mitochondrial Dysfunction: The Road to Alpha-Synuclein Oligomerization in PD. Parkinson's Disease. January 2011, 2011: 693761. PMC 3026982 . PMID 21318163. doi:10.4061/2011/693761.
- ^ Alzheimer’s, Association. 2015 Alzheimer's disease facts and figures.. Alzheimer's & dementia : the journal of the Alzheimer's Association. 2015-03, 11 (3): 332–84 [2020-01-25]. PMID 25984581. doi:10.1016/j.jalz.2015.02.003.
- ^ Dronse, J; Fliessbach, K; Bischof, GN; von Reutern, B; Faber, J; Hammes, J; Kuhnert, G; Neumaier, B; Onur, OA; Kukolja, J; van Eimeren, T; Jessen, F; Fink, GR; Klockgether, T; Drzezga, A. In vivo Patterns of Tau Pathology, Amyloid-β Burden, and Neuronal Dysfunction in Clinical Variants of Alzheimer's Disease.. Journal of Alzheimer's disease : JAD. 2017, 55 (2): 465–471 [2020-01-25]. PMID 27802224. doi:10.3233/JAD-160316.
- ^ Ml, Steinhilb; D, Dias-Santagata; Ta, Fulga; Dl, Felch; Mb, Feany. Tau Phosphorylation Sites Work in Concert to Promote Neurotoxicity in Vivo. Molecular biology of the cell. 2007-12, 18 (12) [2020-01-25]. PMID 17928404 (英语).
- ^ Mocanu, MM; Nissen, A; Eckermann, K; Khlistunova, I; Biernat, J; Drexler, D; Petrova, O; Schönig, K; Bujard, H; Mandelkow, E; Zhou, L; Rune, G; Mandelkow, EM. The potential for beta-structure in the repeat domain of tau protein determines aggregation, synaptic decay, neuronal loss, and coassembly with endogenous Tau in inducible mouse models of tauopathy.. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2008-01-16, 28 (3): 737–48 [2020-01-25]. PMID 18199773. doi:10.1523/JNEUROSCI.2824-07.2008.
- ^ Lee, MJ; Lee, JH; Rubinsztein, DC. Tau degradation: the ubiquitin-proteasome system versus the autophagy-lysosome system.. Progress in neurobiology. 2013-06, 105: 49–59 [2020-01-28]. PMID 23528736. doi:10.1016/j.pneurobio.2013.03.001.
- ^ Chesser, AS; Ganeshan, V; Yang, J; Johnson, GV. Epigallocatechin-3-gallate enhances clearance of phosphorylated tau in primary neurons.. Nutritional neuroscience. 2016, 19 (1): 21–31 [2020-01-28]. PMID 26207957. doi:10.1179/1476830515Y.0000000038.
- ^ Caccamo, A; Magrì, A; Medina, DX; Wisely, EV; López-Aranda, MF; Silva, AJ; Oddo, S. mTOR regulates tau phosphorylation and degradation: implications for Alzheimer's disease and other tauopathies.. Aging cell. 2013-06, 12 (3): 370–80 [2020-01-28]. PMID 23425014. doi:10.1111/acel.12057.
- ^ Cherra SJ, 3rd; Chu, CT. Autophagy in neuroprotection and neurodegeneration: A question of balance.. Future neurology. 2008-05, 3 (3): 309–323 [2020-01-28]. PMID 18806889. doi:10.2217/14796708.3.3.309.
- ^ Wang, Y; Xu, K; Zhang, H; Zhao, J; Zhu, X; Wang, Y; Wu, R. Retinal ganglion cell death is triggered by paraptosis via reactive oxygen species production: a brief literature review presenting a novel hypothesis in glaucoma pathology.. Molecular medicine reports. 2014-09, 10 (3): 1179–83 [2020-01-25]. PMID 24969312. doi:10.3892/mmr.2014.2346.
- ^ Rodríguez-Muela, N; Germain, F; Mariño, G; Fitze, PS; Boya, P. Autophagy promotes survival of retinal ganglion cells after optic nerve axotomy in mice.. Cell death and differentiation. 2012-01, 19 (1): 162–9 [2020-01-25]. PMID 21701497. doi:10.1038/cdd.2011.88.
- ^ Randow, F; Münz, C. Autophagy in the regulation of pathogen replication and adaptive immunity.. Trends in immunology. 2012-10, 33 (10): 475–87 [2020-01-27]. PMID 22796170. doi:10.1016/j.it.2012.06.003.
- ^ Pei, J; Zhao, M; Ye, Z; Gou, H; Wang, J; Yi, L; Dong, X; Liu, W; Luo, Y; Liao, M; Chen, J. Autophagy enhances the replication of classical swine fever virus in vitro.. Autophagy. 2014-01, 10 (1): 93–110 [2020-01-27]. PMID 24262968. doi:10.4161/auto.26843.
- ^ Wang, G; Yu, Y; Tu, Y; Tong, J; Liu, Y; Zhang, C; Chang, Y; Wang, S; Jiang, C; Zhou, EM; Cai, X. Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Infection Induced Apoptosis and Autophagy in Thymi of Infected Piglets.. PloS one. 2015, 10 (6): e0128292 [2020-01-27]. PMID 26046751. doi:10.1371/journal.pone.0128292.
- ^ Meng, C; Zhou, Z; Jiang, K; Yu, S; Jia, L; Wu, Y; Liu, Y; Meng, S; Ding, C. Newcastle disease virus triggers autophagy in U251 glioma cells to enhance virus replication.. Archives of virology. 2012-06, 157 (6): 1011–8 [2020-01-27]. PMID 22398914. doi:10.1007/s00705-012-1270-6.
- ^ Datan, E; Roy, SG; Germain, G; Zali, N; McLean, JE; Golshan, G; Harbajan, S; Lockshin, RA; Zakeri, Z. Dengue-induced autophagy, virus replication and protection from cell death require ER stress (PERK) pathway activation.. Cell death & disease. 2016-03-03, 7: e2127 [2020-01-27]. PMID 26938301. doi:10.1038/cddis.2015.409.
- ^ Sir, D; Tian, Y; Chen, WL; Ann, DK; Yen, TS; Ou, JH. The early autophagic pathway is activated by hepatitis B virus and required for viral DNA replication.. Proceedings of the National Academy of Sciences of the United States of America. 2010-03-02, 107 (9): 4383–8 [2020-01-27]. PMID 20142477. doi:10.1073/pnas.0911373107.
- ^ Ke, PY; Chen, SS. Activation of the unfolded protein response and autophagy after hepatitis C virus infection suppresses innate antiviral immunity in vitro.. The Journal of clinical investigation. 2011-01, 121 (1): 37–56 [2020-01-27]. PMID 21135505. doi:10.1172/JCI41474.
- ^ Levine, B; Mizushima, N; Virgin, HW. Autophagy in immunity and inflammation.. Nature. 2011-01-20, 469 (7330): 323–35 [2020-01-27]. PMID 21248839. doi:10.1038/nature09782.
- ^ Tang, SW; Chen, CY; Klase, Z; Zane, L; Jeang, KT. The cellular autophagy pathway modulates human T-cell leukemia virus type 1 replication.. Journal of virology. 2013-02, 87 (3): 1699–707. PMID 23175371. doi:10.1128/JVI.02147-12.
- ^ Ren, T; Takahashi, Y; Liu, X; Loughran, TP; Sun, SC; Wang, HG; Cheng, H. HTLV-1 Tax deregulates autophagy by recruiting autophagic molecules into lipid raft microdomains.. Oncogene. 2015-01-15, 34 (3): 334–45 [2020-01-27]. PMID 24362528. doi:10.1038/onc.2013.552.
- ^ Deretic, V; Levine, B. Autophagy balances inflammation in innate immunity.. Autophagy. 2018, 14 (2): 243–251 [2020-01-27]. PMID 29165043. doi:10.1080/15548627.2017.1402992.
- ^ Brass, AL; Dykxhoorn, DM; Benita, Y; Yan, N; Engelman, A; Xavier, RJ; Lieberman, J; Elledge, SJ. Identification of host proteins required for HIV infection through a functional genomic screen.. Science (New York, N.Y.). 2008-02-15, 319 (5865): 921–6 [2020-01-27]. PMID 18187620. doi:10.1126/science.1152725.
- ^ Espert, L; Denizot, M; Grimaldi, M; Robert-Hebmann, V; Gay, B; Varbanov, M; Codogno, P; Biard-Piechaczyk, M. Autophagy is involved in T cell death after binding of HIV-1 envelope proteins to CXCR4.. The Journal of clinical investigation. 2006-08, 116 (8): 2161–72 [2020-01-27]. PMID 16886061. doi:10.1172/JCI26185.
- ^ Sagnier, S; Daussy, CF; Borel, S; Robert-Hebmann, V; Faure, M; Blanchet, FP; Beaumelle, B; Biard-Piechaczyk, M; Espert, L. Autophagy restricts HIV-1 infection by selectively degrading Tat in CD4+ T lymphocytes.. Journal of virology. 2015-01, 89 (1): 615–25. PMID 25339774. doi:10.1128/JVI.02174-14.
- ^ Campbell, GR; Rawat, P; Bruckman, RS; Spector, SA. Human Immunodeficiency Virus Type 1 Nef Inhibits Autophagy through Transcription Factor EB Sequestration.. PLoS pathogens. 2015-06, 11 (6): e1005018 [2020-01-27]. PMID 26115100. doi:10.1371/journal.ppat.1005018.
- ^ Zughaier, SM; Kandler, JL; Balthazar, JT; Shafer, WM. Phosphoethanolamine Modification of Neisseria gonorrhoeae Lipid A Reduces Autophagy Flux in Macrophages.. PloS one. 2015, 10 (12): e0144347 [2020-01-27]. PMID 26641098. doi:10.1371/journal.pone.0144347.
- ^ Kim, WJ; Mai, A; Weyand, NJ; Rendón, MA; Van Doorslaer, K; So, M. Neisseria gonorrhoeae evades autophagic killing by downregulating CD46-cyt1 and remodeling lysosomes.. PLoS pathogens. 2019-02, 15 (2): e1007495 [2020-01-27]. PMID 30753248. doi:10.1371/journal.ppat.1007495.
- ^ Lu, P; Wang, S; Lu, Y; Neculai, D; Sun, Q; van der Veen, S. A Subpopulation of Intracellular Neisseria gonorrhoeae Escapes Autophagy-Mediated Killing Inside Epithelial Cells.. The Journal of infectious diseases. 2019-01-01, 219 (1): 133–144 [2020-01-27]. PMID 29688440. doi:10.1093/infdis/jiy237.
- ^ Al-Younes, HM; Brinkmann, V; Meyer, TF. Interaction of Chlamydia trachomatis serovar L2 with the host autophagic pathway.. Infection and immunity. 2004-08, 72 (8): 4751–62 [2020-01-27]. PMID 15271937. doi:10.1128/IAI.72.8.4751-4762.2004.
- ^ Fang, W; Shu, S; Yongmei, L; Endong, Z; Lirong, Y; Bei, S. miR-224-3p inhibits autophagy in cervical cancer cells by targeting FIP200.. Scientific reports. 2016-09-12, 6: 33229. PMID 27615604. doi:10.1038/srep33229.
- ^ Tegelenbosch, RA; de Rooij, DG. A quantitative study of spermatogonial multiplication and stem cell renewal in the C3H/101 F1 hybrid mouse.. Mutation research. 1993-12, 290 (2): 193–200 [2020-01-23]. PMID 7694110. doi:10.1016/0027-5107(93)90159-d.
- ^ Liu, ML; Wang, JL; Wei, J; Xu, LL; Yu, M; Liu, XM; Ruan, WL; Chen, JX. Tri-ortho-cresyl phosphate induces autophagy of rat spermatogonial stem cells.. Reproduction (Cambridge, England). 2015-02, 149 (2): 163–70 [2020-01-23]. PMID 25385720. doi:10.1530/REP-14-0446.
- ^ Xu, LL; Liu, ML; Wang, JL; Yu, M; Chen, JX. Saligenin cyclic-o-tolyl phosphate (SCOTP) induces autophagy of rat spermatogonial stem cells.. Reproductive toxicology (Elmsford, N.Y.). 2016-04, 60: 62–8 [2020-01-23]. PMID 26815770. doi:10.1016/j.reprotox.2016.01.004.
- ^ McNally, K; Berg, E; Cortes, DB; Hernandez, V; Mains, PE; McNally, FJ. Katanin maintains meiotic metaphase chromosome alignment and spindle structure in vivo and has multiple effects on microtubules in vitro.. Molecular biology of the cell. 2014-04, 25 (7): 1037–49 [2020-01-23]. PMID 24501424. doi:10.1091/mbc.E13-12-0764.
- ^ Kabeya, Y; Mizushima, N; Ueno, T; Yamamoto, A; Kirisako, T; Noda, T; Kominami, E; Ohsumi, Y; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing.. The EMBO journal. 2000-11-01, 19 (21): 5720–8 [2020-01-23]. PMID 11060023. doi:10.1093/emboj/19.21.5720.
- ^ Gallardo Bolaños, JM; Miró Morán, Á; Balao da Silva, CM; Morillo Rodríguez, A; Plaza Dávila, M; Aparicio, IM; Tapia, JA; Ortega Ferrusola, C; Peña, FJ. Autophagy and apoptosis have a role in the survival or death of stallion spermatozoa during conservation in refrigeration.. PloS one. 2012, 7 (1): e30688 [2020-01-23]. PMID 22292020. doi:10.1371/journal.pone.0030688.
- ^ Zhuo, C; Ji, Y; Chen, Z; Kitazato, K; Xiang, Y; Zhong, M; Wang, Q; Pei, Y; Ju, H; Wang, Y. Proteomics analysis of autophagy-deficient Atg7-/- MEFs reveals a close relationship between F-actin and autophagy.. Biochemical and biophysical research communications. 2013-08-02, 437 (3): 482–8 [2020-01-23]. PMID 23850690. doi:10.1016/j.bbrc.2013.06.111.
- ^ Mu, Y; Yan, WJ; Yin, TL; Zhang, Y; Li, J; Yang, J. Diet-induced obesity impairs spermatogenesis: a potential role for autophagy.. Scientific reports. 2017-03-09, 7: 43475 [2020-01-23]. PMID 28276438. doi:10.1038/srep43475.
- ^ Abelaira, HM; Réus, GZ; Neotti, MV; Quevedo, J. The role of mTOR in depression and antidepressant responses.. Life sciences. 2014-04-17, 101 (1-2): 10–4 [2020-01-24]. PMID 24582593. doi:10.1016/j.lfs.2014.02.014.
- ^ Polajnar, M; Zerovnik, E. Impaired autophagy: a link between neurodegenerative and neuropsychiatric diseases.. Journal of cellular and molecular medicine. 2014-09, 18 (9): 1705–11 [2020-01-24]. PMID 25139375. doi:10.1111/jcmm.12349.
- ^ Atkin, TA; Brandon, NJ; Kittler, JT. Disrupted in Schizophrenia 1 forms pathological aggresomes that disrupt its function in intracellular transport.. Human molecular genetics. 2012-05-01, 21 (9): 2017–28 [2020-01-24]. PMID 22291444. doi:10.1093/hmg/dds018.
- ^ Machado-Vieira, R; Zanetti, MV; Teixeira, AL; Uno, M; Valiengo, LL; Soeiro-de-Souza, MG; Oba-Shinjo, SM; de Sousa, RT; Zarate CA, Jr; Gattaz, WF; Marie, SK. Decreased AKT1/mTOR pathway mRNA expression in short-term bipolar disorder.. European neuropsychopharmacology : the journal of the European College of Neuropsychopharmacology. 2015-04, 25 (4): 468–73 [2020-01-24]. PMID 25726893. doi:10.1016/j.euroneuro.2015.02.002.
- ^ Kim, HW; Rapoport, SI; Rao, JS. Altered expression of apoptotic factors and synaptic markers in postmortem brain from bipolar disorder patients.. Neurobiology of disease. 2010-03, 37 (3): 596–603 [2020-01-24]. PMID 19945534. doi:10.1016/j.nbd.2009.11.010.
- ^ Son, JH; Shim, JH; Kim, KH; Ha, JY; Han, JY. Neuronal autophagy and neurodegenerative diseases.. Experimental & molecular medicine. 2012-02-29, 44 (2): 89–98 [2020-01-24]. PMID 22257884. doi:10.3858/emm.2012.44.2.031.
- ^ Shen, W; Ganetzky, B. Autophagy promotes synapse development in Drosophila.. The Journal of cell biology. 2009-10-05, 187 (1): 71–9 [2020-01-24]. PMID 19786572. doi:10.1083/jcb.200907109.
- ^ Moosavi, MA; Haghi, A; Rahmati, M; Taniguchi, H; Mocan, A; Echeverría, J; Gupta, VK; Tzvetkov, NT; Atanasov, AG. Phytochemicals as potent modulators of autophagy for cancer therapy.. Cancer letters. 2018-06-28, 424: 46–69 [2020-01-21]. PMID 29474859. doi:10.1016/j.canlet.2018.02.030.
- ^ Ylä-Anttila, P; Vihinen, H; Jokitalo, E; Eskelinen, EL. Monitoring autophagy by electron microscopy in Mammalian cells.. Methods in enzymology. 2009, 452: 143–64 [2020-01-24]. PMID 19200881. doi:10.1016/S0076-6879(08)03610-0.
- ^ Eng, KE; Panas, MD; Karlsson Hedestam, GB; McInerney, GM. A novel quantitative flow cytometry-based assay for autophagy.. Autophagy. 2010-07, 6 (5): 634–41 [2020-01-24]. PMID 20458170. doi:10.4161/auto.6.5.12112.
- ^ Li, Z; Ji, X; Wang, W; Liu, J; Liang, X; Wu, H; Liu, J; Eggert, US; Liu, Q; Zhang, X. Ammonia Induces Autophagy through Dopamine Receptor D3 and MTOR.. PloS one. 2016, 11 (4): e0153526 [2020-01-24]. PMID 27077655. doi:10.1371/journal.pone.0153526.
外部鏈接
- (英文)Autophagy, a journal produced by Landes Bioscience and edited by DJ Klionsky
- (英文)LongevityMeme entry describing PubMed article on the effects of autophagy and lifespan
- (英文)Autophagolysosome on Drugs.com
- (英文)HADb, a Human Autophagy dedicated Database
- (英文)Autophagy DB, an autophagy database that covers all eukaryotes
- (英文)Self-Destructive Behavior in Cells May Hold Key to a Longer Life
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||